Senuweestelsel
| Senuweestelsel | ||
| Inligting en eksterne bronne | ||

| ||
| Die menslike senuweestelsel. | ||
| Inligting | ||
| Latyn | Systema nervosum | |
| Eksterne bronne | ||
| MeSH | D009420 | |
| FMA | 7157 | |
Die senuweestelsel is die deel van ’n dier wat al sy aksies koördineer deur die oordrag van seine na en van verskillende dele van die liggaam. Dié aksies sluit in die beweging van sy spiere en sy sintuiglike prikkels, asook by hoër diersoorte hul emosionele en kognitiewe prosesse. Die senuweestelsel wissel van eenvoudig by die lae diersoorte tot hoogs ingewikkeld by die hoë dieresoorte soos die mens.
Senuweefsel het die eerste keer in wurmagtige organismes ontstaan, sowat 550 tot 600 miljoen jaar gelede.
By gewerweldes bestaan die senuweestelsel uit twee hoofdele, die sentrale en die perifere senuweestelsel. Eersgenoemde bestaan uit die brein en rugmurg en laasgenoemde hoofsaaklik uit senuwees, kabelagtige bondels aksons, of senuvesels, wat die sentrale senuweestelsel met die res van die liggaam verbind. Senuwees wat seine van die brein af stuur, word motoriese of afvoerende senuwees genoem, terwyl dié wat seine van die liggaam na die sentrale stelsel stuur, sintuig- of toevoerende senuwees heet. Rugmurgsenuwees doen albei en word gemengde senuwees genoem.
Die perifere senuweestelsel word verdeel in a) ’n somatiese en b) ’n outonome senuweestelsel en c) die ingewandsenuweestelsel. Somatiese senuwees bemiddel willekeurige beweging. Die outonome senuweestelsel is verder verdeel in die simpatiese en die parasimpatiese senuweestelsel. Die simpatiese stelsel word in tye van nood in werking gestel sodat die liggaam op ’n prikkel kan reageer, terwyl die parasimpatiese stelsel werk wanneer die organisme in ’n ontspanne toestand is. Die ingewandsenuwees beheer die maagdermstelsel. Die outonome en ingewandsenustelsel werk albei onwillekeurig.
Senuwees wat uit die kop kom, word die harsingsenuwees genoem en dié wat uit die rugmurg kom die rugmurgsenuwees.
Die senuweestelsel word beheer deur ’n spesiale soort sel, die neuron, ook bekend as ’n senuweesel. Neurone is so ontwerp dat hulle seine vinnig en presies na ander selle kan stuur. Die seine word gestuur in die vorm van elektrochemiese golwe wat met aksons langs beweeg; laasgenoemde veroorsaak dat chemiese stowwe, of senuoordraers, afgeskei word by skakelplekke, of sinapse. ’n Sel wat ’n sinapssein van ’n neuron ontvang, kan opgewek, geïnhibeer of op ’n ander manier gemoduleer word. Die verbindings tussen neurone kan senunetwerke vorm wat ’n organisme se waarneming van die wêreld en sy optrede bepaal. Benewens neurone bevat die senuweestelsel ander gespesialiseerde selle, wat gliaselle genoem word; hulle verleen strukturele en metaboliese steun.
Senuweestelsels word in die meeste meersellige diere aangetref, maar die ingewikkeldheid daarvan wissel in ’n groot mate.[1] Die enigste meersellige diere wat nie ’n senuweestelsel het nie, is sponsdiertjies, Placozoa en Mesozoa, wat baie eenvoudige liggaamsplanne het. Die senuweestelsel van die radiaal simmetriese organismes ribkwalle en neteldiere (wat seeanemone, korale en jellievisse insluit) bestaan uit ’n verspreide senuweenet. Alle ander dierespesies, buiten ’n paar soorte wurms, het ’n senuweestelsel wat bestaan uit ’n brein, ’n sentrale string of twee en senuwees wat uit die brein en sentrale string uitloop. Die grootte wissel van ’n paar honderd selle in die eenvoudigste wurms tot sowat 300 miljard selle in Afrika-olifante.[2]
Die sentrale senuweestelsel stuur seine van een sel na ’n ander, van een deel van die liggaam na ’n ander en om terugvoer te kry. ’n Wanfunksie in die senuweestelsel kan voorkom weens genetiese afwykings, fisieke skade weens trauma of vergifting, infeksies of bloot ’n hoë ouderdom. Op die vakgebied neurologie word ondersoek ingestel na afwykings van die senuweestelsel en gesoek na maniere om dit te behandel of verhoed. In die perifere senuweestelsel is die mees algemene probleem die gebrek aan senuweegeleiding, wat die gevolg van verskeie probleme is soos diabetiese neuropatie en meervoudige sklerose. Neurowetenskap is die wetenskapsterrein wat fokus op die bestudering van die senuweestelsel.

Struktuur
[wysig | wysig bron]Die senuweestelsel kry sy naam van die woord "senuwees", wat silindriese bondels vesel is wat van die brein en rugmurg af versprei en herhaaldelik vertak na elke deel van die liggaam.[3] Die antieke Egiptenare, Grieke en Romeine het reeds van senuwees geweet,[4] maar die interne struktuur daarvan is nie verstaan totdat die mens dit met ’n mikroskoop kon ondersoek nie.[5]
Mense het tot omstreeks 1900 nog nie geweet neurone is die basiese eenhede van die brein nie (uitgevind deur die Spaanse neurowetenskaplike Santiago Ramón y Cajal). Of tot 1930 dat chemiese oordrag in die brein plaasvind nie (Henry Hallett Dale en Otto Loewi). Ons het begin om die basiese elektriese verskynsel wat neurone gebruik om te kommunikeer, die aksiepotensiaal, in die 1950's te verstaan (Alan Lloyd Hodgkin, Andrew Huxley en John Eccles). In die 1960's het ons bewus geraak van hoe basiese neuronnetwerk-kodestimulusse en dus basiese begrippe moontlik is (David H. Hubel en Torsten Wiesel). Die molekulêre revolusie het in die 1980's deur Amerikaanse universiteite getrek. In die 1990's het die molekulêre meganismes van gedragsverskynsels algemeen bekend geraak (Eric Richard Kandel)."[6] ’n Mikroskopiese ondersoek wys senuwees bestaan hoofsaaklik uit aksons en verskillende membrane wat hulle omring en in bondels verdeel. Die neurone wat tot die senuwees aanleiding gee, lê nie geheel en al binne die senuwees self nie – hul selliggame lê in die brein, die rugmurg en perifere senuweeknope.[3]
Alle diere wat beter as sponse ontwikkel is, het ’n senuweestelsel. Selfs sponse, eensellige diertjies, het ’n meganisme van sel-tot-selseine wat die voorlopers van neurone is.[7] In radiaal simmetriese diere soos die jellievis bestaan die senuweestelsel uit ’n senuweenet, ’n verspreide netwerk van geïsoleerde selle.[8] In Bilateria, wat die grootste deel van bestaande spesies uitmaak, het die senuweestelsel ’n algemene struktuur wat vroeg in die Ediacarium, meer as 550 miljoen jaar gelede, ontstaan het.[9][10]
Selle
[wysig | wysig bron]Die senuweestelsel bevat twee hoofsoorte selle: neurone en glia.
Neurone
[wysig | wysig bron]
Die senuweestelsel word beheer deur ’n spesiale soort sel, die neuron (ook senuselle genoem).[3] Neurone kan op verskeie maniere van ander selle onderskei word, maar hul mees basiese eienskap is dat hulle met ander selle kommunikeer deur middel van sinapse, wat skakelplekke is met die molekulêre vermoë wat die vinnige stuur van seine moontlik maak, óf elektries óf chemies.[3] Baie soorte neurone het ’n akson, ’n protoplasmiese uitloper wat verafgeleë dele van die liggaam kan bereik en duisende sinaptiese kontakte kan maak.[11] Aksons beweeg dikwels deur die liggaam in bondels wat senuwees genoem word.
Selfs in die senuweestelsel van ’n enkele spesie, soos die mens, kan honderde verskillende soorte neurone bestaan, met ’n groot verskeidenheid vorme en funksies.[11] Dit sluit in sintuigsenuwees wat stimulusse soos lig en klank in senuseine omskakel, en motoriese neurone wat senuseine in die aktivering van die spiere of kliere omskakel. In baie spesies neem die meerderheid van neurone egter deel in die vorming van gesentraliseerde strukture (die brein en senuweeknope); hulle ontvang al hul toevoer van ander neurone en stuur hul afvoer na ander neurone.[3]
Glia
[wysig | wysig bron]Gliaselle (van die Griekse naam vir "gom") is niesenuselle wat steun en voeding verskaf, homeostase handhaaf, miëlien vorm en aan seinsendings in die senuweestelsel deelneem.[12] Daar word geraam die getal glia in die menslike brein is omtrent dieselfde as die getal neurone, hoewel die verhoudings in verskillende dele van die brein verskil.[13] Van die belangrikste funksies van gliaselle is om neurone te ondersteun en hulle in plek te hou; voedingstowwe aan neurone te verskaf; neurone elektries te isoleer; patogene te vernietig, dooie neurone te verwyder en die aksons na hul teikens te lei.[12] ’n Baie belangrike soort glia (oligodendrosiete in die sentrale senuweestelsel en Schwannselle in die perifere senuweestelsel) genereer lae van ’n vetterige stof, miëlien, wat om aksons vou en elektriese isolasie verskaf wat hulle toelaat om aksiepotensiale baie vinniger en doeltreffender te stuur.
Anatomie in gewerweldes
[wysig | wysig bron]
Die senuweestelsel van gewerweldes, onder andere mense, is verdeel in die sentrale en die perifere senuweestelsel.[14]
Die sentrale stelsel (SSS) is die grootste en bestaan uit die brein en rugmurg.[14] Die rugmurgkanaal bevat die rugmurg en die harsingsholte die brein. Die stelsel word omring en beskerm deur die meninges, drie vlieslae wat ’n harde, leeragtige buitenste laag, die dura mater, insluit. Die brein word ook deur die skedel beskerm en die rugmurg deur die werwels.
Perifere senuweestelsel (PSS) is ’n versamelnaam vir die strukture van die senuweestelsel wat nie in die SSS geleë is nie.[15] Die meerderheid van die aksonbondels, wat senuwees genoem word, word beskou as deel van die PSS, selfs wanneer die selliggaam van die neuron waartoe hulle behoort, in die brein of rugmurg lê.
Die perifere senuweestelsel word verdeel in die somatiese en die outonome of ingewandsdeel. Die somatiese deel bestaan uit die senuwees in die vel, gewrigte en spiere. Die selliggaam van somatiese sintuigneurone lê in die dorsale wortelganglia van die rugmurg. Die ingewandsdeel bevat neurone wat die interne organe, bloedvate en kliere besenu. Die ingewandsdeel bestaan self uit twee dele: die simpatiese en parasimpatiese senuweestelsel. Sommige skrywers sluit ook die sintuigneurone waarvan die selliggame in die ledemate lê (vir sintuie soos gevoel) in as deel van die PSS, maar nie almal nie.[16]
Gewerweldes se senuweestelsel kan ook verdeel word in streke wat grysstof en witstof genoem word.[17] Grysstof, wat net grys is in gepreserveerde weefsel maar in lewende weefsel eerder pienk of ligbruin is, bevat ’n groot proporsie van selliggame van neurone. Witstof bestaan hoofsaaklik uit gemiëlineerde aksons en kry sy kleur van die miëlien. Dit sluit in al die senuwees en ’n groot deel van die binnekant van die brein en rugmurg. Grysstof word aangetref in bondels van neurone in die brein en rugmurg, en in skorslae op hul oppervlak. Daar is ’n anatomiese konvensie dat ’n bondel neurone in die brein of rugmurg ’n kern genoem word, terwyl dit in die ledemate ’n ganglion is. Daar is egter uitsonderings, veral in die deel van die voorbrein wat die "basale ganglia" genoem word.[18]
Vergelykende anatomie en evolusie
[wysig | wysig bron]Voorlopers van neurone in sponse
[wysig | wysig bron]Sponsdiertjies het geen selle wat deur sinapse aan mekaar verbind is nie, en dus geen neurone en geen senuweestelsel nie. Hulle het egter homoloë van baie gene wat sleutelrolle speel in sinaptiese funksie. Onlangse studies het gewys sponsselle toon ’n groep proteïene wat saambondel om ’n struktuur te vorm wat lyk na ’n postsinaptiese digtheid (die seinontvangende deel van ’n sinaps).[7] Die funksie van hierdie struktuur is egter tans onduidelik. Hoewel sponsselle nie sinaptiese transmissies toon nie, kommunikeer hulle wel met mekaar deur kalsiumgolwe en ander impulse, wat sommige eenvoudige aksies soos liggaamsametrekkings bemiddel.[19]
Radiata
[wysig | wysig bron]Radiata (radiaal simmetriese diere) soos neteldiere, ribkwalle en verwante diere het verspreide senuweenetwerke eerder as ’n sentrale senuweestelsel. In die meeste neteldiere is die netwerk min of meer egalig deur die liggaam versprei; in ribkwalle is dit naby die mond gekonsentreer. Die senuweenetwerke bestaan uit sintuigneurone, wat chemiese, tas- en visuele seine opvang; motoriese neurone, wat sametrekkings van die liggaamswand kan aktiveer; en intermediêre neurone, wat patrone aktiwiteite in die sintuigneurone bespeur en in antwoord daarop seine stuur na groepe motororiese neurone. In sommige gevalle is groepe intermediêre neurone in diskrete ganglia gebondel.[8]
Die ontwikkeling van die senuweestelsel in Radiata is relatief ongestruktureer. Anders as Bilateria (bilateraal simmetriese diere) het Radiata net twee basiese kiemlae, die endoderm en ektoderm. Neurone word gegenereer vanuit ’n spesiale stel ektodermiese voorloperselle, wat ook dien as voorlopers van elke ander ektodermiese seltipe.[20]
Bilateria
[wysig | wysig bron]
Die oorgrote meerderheid bestaande diere is Bilateria, wat beteken hul linker- en regterkant is min of meer spieëlbeelde van mekaar. Alle Bilateria stam vermoedelik af van ’n gemeenskaplike wurmagtige voorsaat wat in die Ediacarium-periode, sowat 550 miljoen tot 600 miljoen jaar gelede, voorgekom het.[9] Die basiese Bilateria-liggaamsvorm is ’n buis met ’n hol ingewandsholte wat van die mond na die anus loop, ’n senuweestring met ’n vergroting ("ganglion") vir elke liggaamsegment en ’n ekstragroot ganglion aan die voorkant, die "brein".
Selfs soogdiere, insluitende die mens, toon die gesegmenteerde bilaterale liggaamsplan op die vlak van die senuweestelsel. Die rugmurg bevat ’n reeks gesegmenteerde ganglia, wat elk aanleiding gee tot motoriese en sintuigsenuwees wat ’n deel van die liggaamsoppervlak en onderliggende spierstelsel besenu. In die ledemate is die uitleg van die besenu-patroon ingewikkeld, maar in die romp gee dit aanleiding tot ’n reeks nou bande. Die topdriesegmente behoort tot die brein en gee aanleiding tot die voor-, middel- en agterbrein.[21]
Bilateria kan, geskoei op gebeure baie vroeg in embrio-ontwikkeling, in twee superfilums verdeel word: Protostomia en Deuterostomia.[22] Laasgenoemde sluit in gewerweldes sowel as stekelhuidiges, Hemichordata en Xenoturbellida.[23] Protostomia, die meer uiteenlopende groep, sluit in geleedpotiges, weekdiere en verskeie ander soorte wurms. Daar is ’n basiese verskil tusen die twee groepe in die plasing van die senuweestelsel in die liggaam: Protostomia het ’n senuweestring in die ventrale (gewoonlik onderste) kant van die liggaam, terwyl die senuweestring by Deuterostomia in die dorsale (gewoonlik boonste) kant lê. Verskeie aspekte van die liggaam is eintlik omgekeerd in die twee groepe, insluitend die uitdrukkingspatrone van verskeie gene wat dorsaal-na-ventraal-gradiënte toon. Die meeste anatome glo nou die liggaam van die twee groepe is teenoorgesteldes van mekaar, ’n hipotese wat die eerste keer deur Geoffroy Saint-Hilaire voorgestel is vir insekte in vergelyking met gewerweldes.[24]
Wurms
[wysig | wysig bron]
Wurms is die eenvoudigste bilateraal simmetriese diere en toon ’n basiese struktuur van die bilaterale senuweestelsel. Erdwurms het byvoorbeeld ’n dubbele senuweestring wat met hul hele lyf langs loop en by die stert en mond byeenkom. Dié senuweestringe is oor die dwarste verbind deur senuwees, soos die trappies van ’n leer. Die dwarsgeleë senuwees help die twee kante van die dier koördineer. Twee ganglia aan die kopkant werk soos ’n eenvoudige brein. Eenvoudige oë verskaf inligting oor lig en donkerte.[25]
Geleedpotiges
[wysig | wysig bron]
Geleedpotiges soos insekte en skaaldiere se senuweestelsel bestaan uit ’n reeks ganglia, wat deur ’n ventrale senuweestring verbind word; dié bestaan uit twee parallelle dele wat met die lengte van die abdomen langs loop.[26] Elke liggaamsegment het ’n ganglion aan elke kant, maar sommige ganglia is saamgesmelt om die brein en ander groot ganglia te vorm. Die kopsegment bevat die brein. Die brein is verdeel in drie dele. Net daaragter lê die benedeslukderm-ganglion, wat uit drie pare saamgesmelte ganglia bestaan. Dit beheer die monddele, speekselkliere en sekere spiere. Baie geleedpotiges het goed ontwikkelde sintuigorgane, insluitend saamgestelde oë vir sig en antennes vir reuk en feromoonsensasie. Die inligting van hierdie organe af word deur die brein verwerk.
Baie neurone by insekte het selligame wat aan die kant van die brein geleë en elektries passief is – hulle verskaf net metaboliese steun en stuur nie seine nie. ’n Protoplasmiese vesel loop uit die selliggaam en vertak; sekere dele stuur seine en ander ontvang dit. Die meeste dele van die insektebrein het dus passiewe selliggame wat om die buitekant gerangskik is, terwyl die senuseinverwerking in die agterkant plaasvind in ’n netwerk protoplasmiese vesels wat senupole genoem word.[27]
Funksie
[wysig | wysig bron]Op die mees basiese vlak is die funksie van die senuweestelsel om seine van een sel na ’n ander te stuur, of van een deel van die liggaam na ’n ander. Daar is talle maniere waarop ’n sel seine na ander selle kan stuur. Een is deur chemiese stowwe, genaamd hormone, na die interne sirkulasie te stuur sodat hulle na verafgeleë plekke kan versprei. In teenstelling met hierdie "uitsaaimodus" van seine, maak die senuweestelsel punt-tot-punt-seine moontlik – neurone projekteer hul aksons na spesifieke teikengebiede en maak sinaptiese verbindings met spesifieke teikenselle.[28]
Op ’n meer integrerende vlak is die hooffunksie van die senuweestelsel om die liggaam te beheer.[3] Dit word gedoen deur die verkryging van inligting uit die omgewing deur middel van sintuigreseptors, die stuur van seine wat hierdie inligting in die sentrale senuweestelsel enkodeer, die verwerking van die inligting om ’n geskikte reaksie te bepaal en die stuur van seine na spiere of kliere om die gewenste reaksie te aktiveer. Die evolusie van ’n ingewikkelde senuweestelsel het dit vir verskeie dierespesies moontlik gemaak om gevorderde waarnemingsvermoëns soos sig, komplekse sosiale interaksies, die vinnige koördinasie van orgaanstelsels en die geïntegreerde verwerking van gelyktydige seine te hê. By mense stel die sofistikasie van die senuweestelsel taal, die abstrakte voorstelling van begrippe, die uitruiling van kultuur en baie ander eienskappe van menslike gemeenskappe daar wat nie sonder die menslike brein moontlik sou gewees het nie.
Neurone en sinapse
[wysig | wysig bron]Die meeste neurone stuur seine deur hul aksons, hoewel sommige soorte seine van dendriet tot dendriet stuur. Sommige selle het nie aksons nie en kommunikeer slegs deur hul dendriete. Senuweeseine plant deur ’n akson voort in die vorm van elektrochemiese golwe wat aksiepotensiale genoem word; hulle produseer sel-tot-selseine by punte waar aksonterminale sinaptiese kontak met ander selle maak.[29]

Sinapse kan elektries of chemies wees. Elektriese sinapse maak direkte elektriese konneksies tussen neurone,[30] maar chemiese sinapse is baie meer algemeen en baie meer uiteenlopend wat funksie betref.[31] By ’n chemiese sinaps word die sel wat seine stuur presinapties genoem, en die sel wat seine ontvang postsinapties. Beide die presinaptiese en postsinaptiese gebiede is vol molekulêre masjinerie wat die seinproses uitvoer. Die presinaptiese area bevat groot hoeveelhede klein sferiese houers wat sinapsblasies genoem word; hulle is vol senuoordragschemikalieë.[29]
Wanneer die presinaptiese terminaal elektries gestimuleer word, word ’n reeks molekules wat in die membraan lê, geaktiveer en veroorsaak dat die inhoud van die blasie vrygestel word in die smal spasie tussen die presinaptiese en postsinaptiese membraan, wat die sinaptiese spleet genoem word. Die senuoordraer verbind dan met reseptore in die postsinaptiese membraan en veroorsaak dat hulle ’n geaktiveerde toestand binnegaan.[31] Na gelang van die soort reseptor, kan die postsinaptiese sel opgewek, geïnhibeer of op baie ander maniere verander word. Die vrystelling van die senuoordraer asetielcholien by ’n sinaptiese kontak tussen ’n motoriese neuron en ’n spiersel veroorsaak byvoorbeeld die vinnige sametrekking van die spiersel.[32] Die hele sinaptiese oordragsproses neem net ’n breukdeel van ’n millisekonde, hoewel die uitwerking op die postsinaptiese sel baie langer kan duur (selfs oneindig lank in gevalle waar die sinaptiese sein lei tot die vorming van ’n geheuespoor).[11]

Daar is letterlik honderde verskillende soorte sinapse. Eintlik is daar meer as honderd bekende senuoordraers, en baie van hulle het verskeie soorte reseptore.[33] Baie sinapse gebruik meer as een senuoordraer – dit is algemeen dat ’n sinaps een vinnigwerkende kleinmolekule-senuoordraer kan gebruik, soos glutaminesuur of gamma-aminobottersuur, asook een of meer peptied-senuoordraers wat stadiger moduleringsrolle speel.
Volgens ’n reël bekend as die beginsel van Dale, wat slegs ’n paar bekende uitsonderings het, stel ’n neuron dieselfde senuoordaers vry by al sy sinapse.[34] Dit beteken nie die neuron het dieselfde uitwerking op al sy teikens nie – die uitwerking van ’n sinaps hang nie van die senuoordraer af nie, maar van die reseptore wat dit aktiveer.[31] Omdat verskillende teikens verskillende soorte reseptore kan gebruik (en dit dikwels doen), is dit moontlik dat ’n neuron ’n opwekkende uitwerking op een stel teikenselle kan hê, ’n inhiberende uitwerking op ’n tweede stel en ’n komplekse modulerende uitwerking op ’n derde stel. Nietemin het die twee mees algemene senuoordraers, glutaminesuur en gamma-aminobottersuur, albei ’n baie konsekwente uitwerking. Glutaminesuur het verskeie wyd voorkomende soorte reseptore, maar hulle is almal opwekkend of modulerend. Net so het gamma-aminobottersuur verskeie wyd voorkomende reseptortipes, maar hulle is almal inhiberend.
Een baie belangrike subgroep sinapse kan geheuespore vorm deur middel van langdurige aktiwiteitsafhanklike veranderings in sinaptiese sterkte.[35] Die bekendste vorm van neurale geheue is ’n proses wat langtermynversterking genoem word; dit werk by sinapse wat die senuoordraer glutaminesuur gebruik wat werk op ’n spesiale soort reseptor bekend as ’n NMDA-reseptor.[36] Dié reseptor het ’n "assosiatiewe" eienskap: as die twee selle wat in die sinaps betrokke is albei op min of meer dieselfde tyd geaktiveer word, open ’n kanaal wat kalsium toelaat om in die teikensel te vloei.[37] Die kalsium begin ’n tweede boodskapstroom wat eindelik lei tot ’n toename in die getal glutaminesuurreseptore in die teikensel. Daardeur word die effektiewe sterkte van die sinaps verhoog. Hierdie verandering in sterkte kan weke of langer duur. Sedert die ontdekking van langtermynversterking in 1973, is baie ander soorte sinaptiese geheuespore ontdek wat ’n toe- of afname in sinaptiese sterkte behels en wat ’n verskeidenheid tydperke duur.[36] Die beloningstelsel, wat gewenste gedrag aanmoedig, is byvoorbeeld afhanklik van ’n vorm van langtermynversterking waar dopamien as senuoordraer gebruik word.[38] Al hierdie vorme van sinaptiese modifieerbaarheid gee gesamentlik aanleiding tot neurale vormbaarheid, of die vermoë van die senuweestelsel om by veranderings in die omgewing aan te pas.
Senukringlope en stelsels
[wysig | wysig bron]
Die funksie van die senuweestelsel is histories baie jare lank beskou as ’n stimulus-reaksie-verbinding.[39] Volgens dié beskouing begin neurale prosessering met stimulusse wat sensoriese neurone aktiveer en seine vervaardig wat deur kettings verbindings in die rugmurg en brein voortplant; dit lei eindelik tot die aktivering van motororiese neurone en daardeur tot spiersametrekking. René Descartes het geglo elke gedrag van ’n dier en die meeste optredes van mense kan verduidelik word aan die hand van stimulus-reaksie-kringlope. Hy het egter ook geglo hoër kognitiewe funksies soos taal kan nie meganisties verduidelik word nie.[40] Charles Sherrington het in sy invloedryke boek van 1906, The Integrative Action of the Nervous System,[39] die begrip van stimulus-reaksie-meganismes in veel groter besonderhede ontwikkel, en behaviorisme, die denkskool wat die psigologie deur die middel van die 20ste eeu oorheers het, het elke aspek van menslike gedrag in stimulus-reaksie-terme probeer verduidelik.[41]
Studies in elektrofisiologie wat vroeg in die 20ste eeu begin is en teen die 1940's ’n hoogtepunt bereik het, het getoon die senuweestelsel bevat baie meganismes wat self patrone aktiwiteit genereer, sonder ’n stimulus van buite.[42] Daar is bevind neurone is in staat om gereelde reekse aksiepotensiale, of reekse uitbarstings, te produseer – selfs in algehele isolasie.[43]
Wanneer intrinsiek aktiewe neurone in ingewikkelde kringlope aan mekaar verbind word, word die moontlikhede vir die generering van ingewikkelde temporale patrone baie groter.[44] Volgens ’n moderne opvatting word die funksie van die senuweestelsel deels beskou in terme van stimulus-reaksie-kettings, en gedeeltelik in terme van intrinsiek gegenereerde aktiwiteitspatrone – albei soorte aktiwiteite tree in wisselwerking met mekaar om die volle repertoire van gedrag te genereer.[45]
Reflekse en ander stimulus-reaksie-kringlope
[wysig | wysig bron]Die eenvoudigste soort senukringloop is ’n refleksboog; dit begin met ’n sensoriese toevoer en eindig met ’n motoriese afvoer, en gaan deur ’n reeks neurone wat verbind is.[46] Dit kan geïllustreer word deur die "wegrukrefleks", wat veroorsaak dat ’n hand weggeruk word nadat dit aan ’n warm stoof geraak het.

Die kringloop begin met sensoriese reseptore in die vel wat geaktiveer word deur ’n skadelike hittevlak: ’n Spesiale soort molekulêre struktuur in die membraan veroorsaak dat hitte die elektriese veld oor die membraan verander. As die verandering in elektriese potensiaal groot genoeg is om die gegewe grens oor te steek, ontlok dit ’n aksiepotensiaal, wat deur die akson van die reseptorsel tot in die rugmurg gestuur word. Daar maak die akson eksitatoriese (opwekkende) sinaptiese kontakte met ander selle, waarvan sommige projekteer (aksonale afvoer stuur) na dieselfde streek van die rugmurg. Ander projekteer na die brein. Een teiken is ’n stel rugmurg-interneurone wat projekteer na motoriese neurone wat die armspiere beheer. Die interneurone wek die motoriese neurone op en as die opwekking sterk genoeg is, genereer sommige van die motoriese neurone aksiepotensiale, wat met hul aksons af beweeg na die punt waar hulle eksitatoriese sinaptiese kontakte met spierselle maak. Die eksitatoriese seine veroorsaak ’n sametrekking van die spierselle, en die arm word weggeruk.
In werklikheid kan daar verskeie komplikasies in hierdie eenvoudige voorstelling wees.[46] Hoewel daar vir die eenvoudigste reflekse kort neurale paaie is van die sensoriese na die motoriese neurone, is daar ook ander nabygeleë neurone wat aan die kringloop deelneem en die reaksie moduleer. Verder is daar projeksies van die brein na die rugmurg wat die vermoë het om die refleks te vergroot of verklein.
Hoewel die eenvoudigste reflekse bemiddel kan word deur kringlope wat heeltemal binne die rugmurg lê, is meer ingewikkelde reaksies afhanklik van seinprosessering in die brein.[47] ’n Voorbeeld is wanneer iets aan die buitekant van ’n mens se visuele veld beweeg en die mens daarheen kyk. Baie stadiums van seinprosessering word dan geïnisieer. Die aanvanklike sensoriese reaksie, in die retina van die oog, en die finale motoriese reaksie, in die breinstam, is glad nie soveel anders as dié in die eenvoudige refleks nie, maar die tussenstadiums verskil heeltemal. In plaas van een of twee stappe in die ketting van prosessering, beweeg die visuele seine deur miskien tientalle stadiums van integrasie, waarin die talamus, harsingskors, basale ganglia, superieure colliculus, kleinharsings en verskeie breinstamkerns betrek word. Hierdie streke voer seinprosesseringsfunksies uit wat insluit omgewingswaarneming, die ontleding van die waarneming, geheueherroeping, besluitneming en motoriese beplanning.[48]
Omgewingswaarneming is die vermoë om biologies relevante inligting uit ’n kombinasie van sensoriese seine te onttrek.[49] Wat die sigsintuig betref, is sensoriese reseptore in die retina van die oog byvoorbeeld individueel net in staat om "ligpunte" in die buitewêreld waar te neem.[50] Tweedevlak-signeurone ontvang toevoer van groepe primêre reseptore; hoërvlak-neurone ontvang toevoer van groepe tweedevlak-neurone, ens. So vorm hulle ’n hiërargie van prosesseringstadiums. In elke stadium word belangrike inligting uit die sein-ensemble onttrek en onbelangrike inligting verwerp. Teen die einde van die proses is toevoerseine wat "ligpunte" verteenwoordig, omgeskakel in ’n neurale voorstelling van voorwerpe in die omgewing en hul eienskappe. Die mees gesofistikeerde sensoriese prosessering vind in die brein plaas, maar ingewikkelde inligtingsonttrekking vind ook in die rugmurg en in buite- sensoriese organe soos die retina plaas.
Intrinsieke patroonopwekking
[wysig | wysig bron]Hoewel stimulus-reaksie-meganismes die maklikste is om te verstaan, kan die senuweestelsel ook die liggaam op maniere beheer wat nie ’n eksterne stimulus verg nie, deur middel van intern opgewekte ritmes van aktiwiteit. Vanweë die verskeidenheid spanningsensitiewe ioonkanale wat in die membraan van die neuron kan voorkom, is baie soorte neurone in staat om (selfs as hulle in isolasie voorkom) ritmiese reekse aksiepotensiale te genereer, of ritmiese wisselings tussen hoëtempo-uitbarstings en rus. Wanneer neurone wat intrinsiek ritmies is, deur eksitatoriese of inhiberende sinapse aan mekaar verbind is, is die gevolglike netwerke in staat tot ’n uiteenlopende getal dinamiese gedragswyses, insluitende aantrekkingsdinamika, periodisiteit en selfs chaos. ’n Netwerk neurone wat sy interne struktuur gebruik om tydelik gestruktureerde afvoer te genereer, sonder dat dit ’n ooreenstemmende tydelik gestruktureerde stimulus vereis, word ’n sentrale patroonopwekker genoem.
Interne patroonopwekking werk op ’n groot verskeidenheid tydskale, van millisekondes tot ure of langer. Een van die belangrikste soorte slaappatrone is sirkadiese ritme – betreffende biologiese prosesse wat al om die 24 uur plaasvind. Alle diere wat bestudeer is, toon sirkadiese wisselings in neurale aktiwiteit, wat gedrag soos die slaap-wakker-siklus beheer. Eksperimentele studies van die 1990's af wys sirkadiese ritme word opgewek deur ’n "genetiese klok" wat bestaan uit ’n spesiale stel gene waarvan die uitdrukkingsvlak deur die loop van die dag styg en daal. Diere so uiteenlopend soos insekte en gewerweldes deel ’n soortgelyke genetiese ritmieseklokstelsel. Die klok word deur lig beïnvloed, maar hou aan werk al word ligvlakke konstant gehou en al is daar geen ander eksterne aanduidings oor watter tyd van die dag dit is nie. Die klokgene vind uitdrukking in baie dele van die senuweestelsel sowel as baie perifere organe, maar in soogdiere word al hierdie "weefselklokke" gesinkroniseer gehou deur seine wat uitgestuur word deur die meestertydhouer in ’n klein deel van die brein bekend as die suprachiasmatiese kern.
Ontwikkeling
[wysig | wysig bron]In gewerweldes sluit belangrike oomblikke in die neurale ontwikkeling van die embrio die volgende in: die ontstaan en differensiasie van neurone uit stamselvoorlopers, die migrasie van onvolgroeide neurone van hul ontstaanplek in die embrio na hul finale posisie, die uitgroei van aksons uit neurone en die geleiding van die beweeglike groeikeël deur die embrio na postsinaptiese vennote, die voortbrenging van sinapse tussen hierdie aksons en hul postsinaptiese vennote, en eindelik die lewenslange verandering in sinapse wat vermoedelik ten grondslag lê van leer en geheue.[51]
Alle diere van Bilateria vorm vroeg in hul ontwikkeling ’n gastrula, wat gepolariseer is; die een kant word die dierpool genoem en die ander kant die plantpool. Die gastrula het die vorm van ’n skyf met drie lae selle: ’n binneste laag wat die endoderm genoem word en wat aanleiding gee tot die voering van die meeste interne organe, ’n middelste laag, die mesoderm, wat aanleiding gee tot die bene en spiere, en ’n buitenste laag, die ektoderm, wat aanleiding gee tot die vel en senuweestelsel.[52]
 |
 |
In gewerweldes is die eerste teken van die senuweestelsel die verskyning van ’n dun strook selle al langs die middel van die rug wat die senuplaat genoem word. Die binneste deel van die senuplaat (met die middellyn langs) sal eindelik die sentrale senuweestelsel (SSS) word en die buitenste deel die perifere senuweestelsel (PSS). Namate die ontwikkeling vorder, verskyn ’n vou (die senugleuf) langs die middellyn. Dié vou verdiep en die bokant groei toe. In dié stadium verskyn die toekomstige SSS as ’n silindriese struktuur wat die senubuis genoem word, terwyl die toekomstige PSS verskyn as twee stroke weefsel bekend as die senulys; dit loop in die lengte bo die senubuis. Die stadiums van senuplaat tot senubuis en senulys is bekend as neurulasie.
Die aard van die induksieproses was onbekend totdat dit eindelik in die 1990's met ’n genetiese benadering opgelos is. Die induksie van die senuweefsel vereis die inhibering van die geen vir ’n sogenaamde beenmorfogenetiese proteïen, of BMP. Veral die proteïen BMP4 is blykbaar betrokke. Twee proteïene wat albei deur die mesoderm afgeskei word, kan BMP4 inhibeer en daardeur die ektoderm induseer om in senuweefsel om te sit. Dit lyk of dieselfde soort molekulêre meganisme betrokke is by baie uiteenlopende soorte diere, van geleedpotiges tot gewerweldes.
Die induksie van senuweefsel lei tot die vorming van voorlopersenuselle, genaamd neuroblaste.[53] Van daar af verdeel selle om óf neurone óf gliaselle te vorm.
Patologie
[wysig | wysig bron]- Die hoofartikel vir hierdie afdeling is: Neurologie.
Neurologie behels die diagnose en behandeling van alle soorte toestande en siektes wat met die sentrale en die perifere senuweestelsel verband hou.[54]
Die sentrale senuweestelsel word beskerm deur groot fisieke en chemiese versperrings. Fisiek word die brein en rugmurg omring deur taai membrane en die bene van die skedel en rugwerwels, wat saam ’n sterk fisieke skild vorm. Chemies word die brein en rugmurg geïsoleer deur die sogenaamde bloed-brein-versperring, wat die meeste soorte chemiese stowwe verhoed om uit die bloedstroom na die binnekant van die SSS te beweeg. Hierdie beskerming maak die SSS in baie opsigte minder kwesbaar as die PSS; aan die ander kant het skade aan die SSS ernstiger gevolge.
Hoewel senuwees gewoonlik diep onder die vel lê, buiten met uitsonderings soos die elmboogsenuwee, is hulle steeds redelik blootgestel aan fisieke skade, wat pyn en ’n verlies aan sensasie of spierbeheer tot gevolg kan hê. Skade aan senuwees kan ook deur swelling en kneusings veroorsaak word op plekke waar ’n senuwee deur ’n nou, benerige kanaal loop, soos in karpale tonnelsindroom. As ’n senuwee heeltemal deurgesny word, sal dit dikwels degenereer, maar vir lang senuwees sal dit maande duur voor die proses afgehandel is.
Benewens fisieke skade, kan perifere neuropatie veroorsaak word deur baie ander mediese probleme, onder meer genetiese toestande, metaboliese toestande soos diabetes, inflammatoriese toestande soos Guillain-Barré-sindroom, ’n vitamientekort, siektes soos melaatsheid en gordelroos, of vergiftiging deur stowwe soos swaarmetale. Baie gevalle het geen bekende oorsake nie en word "idiopaties" genoem. Senuwees kan ook tydelik hul funksie verloor, wat ’n styfheid veroorsaak. Dit word veral veroorsaak deur meganiese druk, ’n daling in temperatuur of chemiese wisselwerkings met sekere medikasie vir plaaslike verdowing.
Fisieke skade aan die rugmurg kan lei tot verlamming, wat permanent kan wees. Eksperimentele studies het getoon rugsenuvesels probeer teruggroei op dieselfde manier as ander senuvesels, maar in die rugmurg lei die vernietiging van weefsel gewoonlik tot die vervaardiging van letselweefsel waardeur die teruggroeiende senuwees nie kan dring nie.
Sien ook
[wysig | wysig bron]Verwysings
[wysig | wysig bron]- ↑ "Nervous System". Columbia Encyclopedia. Columbia University Press.
- ↑ Herculano-Houzel S, Avelino-de-Souza K, et al. (2014). "The elephant brain in numbers". Front Neuroanat. 8: 46. doi:10.3389/fnana.2014.00046. PMC 4053853. PMID 24971054.
- ↑ 3,0 3,1 3,2 3,3 3,4 3,5 "Ch. 2: Nerve cells and behavior". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ Finger S (2001). "Ch. 1: The brain in antiquity". Origins of neuroscience: a history of explorations into brain function. Oxford Univ. Press. ISBN 978-0-19-514694-3.
- ↑ Finger, pp. 43–50
- ↑ Nikoletseas Michael M. (2010) Behavioral and Neural Plasticity. ISBN 978-1453789452
- ↑ 7,0 7,1 Sakarya O, Armstrong KA, Adamska M, et al. (2007). Vosshall L (red.). "A post-synaptic scaffold at the origin of the animal kingdom". PLoS ONE. 2 (6): e506. doi:10.1371/journal.pone.0000506. PMC 1876816. PMID 17551586.
- ↑ 8,0 8,1 Ruppert E.E., Fox R.S., Barnes R.D. (2004). Invertebrate Zoology (7 uitg.). Brooks / Cole. pp. 111–124. ISBN 0-03-025982-7.
{{cite book}}: AS1-onderhoud: gebruik authors-parameter (link) - ↑ 9,0 9,1 Balavoine G (2003). "The segmented Urbilateria: A testable scenario". Int Comp Biology. 43 (1): 137–47. doi:10.1093/icb/43.1.137.
- ↑ Ortega-Hernandez, Javier (29 Februarie 2016), Our 500 million-year-old nervous system fossil shines a light on animal evolution, The Conversation US, Inc., https://theconversation.com/our-500-million-year-old-nervous-system-fossil-shines-a-light-on-animal-evolution-55460, besoek op 6 Mar 2016
- ↑ 11,0 11,1 11,2 "hoofst. 4: The cytology of neurons". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ 12,0 12,1 Allen N.J., Barres B.A. (2009). "Neuroscience: Glia - more than just brain glue". Nature. 457 (7230): 675–7. doi:10.1038/457675a. PMID 19194443.
{{cite journal}}: AS1-onderhoud: gebruik authors-parameter (link) - ↑ Azevedo F.A., Carvalho L.R., Grinberg L.T. (2009). "Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain". J. Comp. Neurol. 513 (5): 532–41. doi:10.1002/cne.21974. PMID 19226510.
{{cite journal}}: AS1-onderhoud: gebruik authors-parameter (link) - ↑ 14,0 14,1 "Ch. 17: The anatomical organization of the central nervous system". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ Standring, Susan (hoofredakteur) (2005). Gray's Anatomy (39ste uitg.). Elsevier Churchill Livingstone. pp. 233–234. ISBN 978-0-443-07168-3.
- ↑ Hubbard JI (1974). The peripheral nervous system. Plenum Press. p. vii. ISBN 978-0-306-30764-5.
- ↑ Purves D, Augustine GJ, Fitzpatrick D, Hall WC, LaMantia AS, McNamara JO, White LE (2008). Neuroscience. 4de uitg. Sinauer Associates. pp. 15–16.
- ↑ Afifi AK (Julie 1994). "Basal ganglia: functional anatomy and physiology. Part 1". J. Child Neurol. 9 (3): 249–60. doi:10.1177/088307389400900306. PMID 7930403.
- ↑ Jacobs DK, Nakanishi N, Yuan D, Camara A, Nichols SA, Hartenstein V, et al. (2007). "Evolution of sensory structures in basal metazoa". Integr Comp Biol. 47 (5): 712–723. doi:10.1093/icb/icm094. PMID 21669752.
- ↑ Sanes DH, Reh TA, Harris WA (2006). Development of the nervous system. Academic Press. pp. 3–4. ISBN 978-0-12-618621-5.
- ↑ Ghysen A (2003). "The origin and evolution of the nervous system". Int. J. Dev. Biol. 47 (7–8): 555–62. CiteSeerX 10.1.1.511.5106. PMID 14756331.
- ↑ Erwin DH, Davidson EH (Julie 2002). "The last common bilaterian ancestor". Development. 129 (13): 3021–32. PMID 12070079. Geargiveer vanaf die oorspronklike op 14 Oktober 2008. Besoek op 4 Junie 2017.
- ↑ Bourlat SJ, Juliusdottir T, Lowe CJ, et al. (November 2006). "Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida". Nature. 444 (7115): 85–8. doi:10.1038/nature05241. PMID 17051155.
- ↑ Lichtneckert R, Reichert H (Mei 2005). "Insights into the urbilaterian brain: conserved genetic patterning mechanisms in insect and vertebrate brain development". Heredity. 94 (5): 465–77. doi:10.1038/sj.hdy.6800664. PMID 15770230.
- ↑ ADEY WR (Februarie 1951). "The nervous system of the earthworm Megascolex". J. Comp. Neurol. 94 (1): 57–103. doi:10.1002/cne.900940104. PMID 14814220.
- ↑ Chapman RF (1998). "Ch. 20: Nervous system". The insects: structure and function. Cambridge University Press. pp. 533–568. ISBN 978-0-521-57890-5.
- ↑ Chapman, p. 546
- ↑ Gray PO (2006). Psychology (5 uitg.). Macmillan. p. 170. ISBN 978-0-7167-7690-1.
- ↑ 29,0 29,1 "Ch. 9: Propagated signaling: the action potential". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ Hormuzdi SG, Filippov MA, Mitropoulou G, et al. (2004). "Electrical synapses: a dynamic signaling system that shapes the activity of neuronal networks". Biochim. Biophys. Acta. 1662 (1–2): 113–37. doi:10.1016/j.bbamem.2003.10.023. PMID 15033583.
- ↑ 31,0 31,1 31,2 "Ch. 10: Overview of synaptic transmission". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ "Ch. 11: Signaling at the nerve-muscle synapse". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ "Ch. 15: Neurotransmitters". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ Strata P, Harvey R (1999). "Dale's principle". Brain Res. Bull. 50 (5–6): 349–50. doi:10.1016/S0361-9230(99)00100-8. PMID 10643431.
- ↑ Paradiso MA, Bear MF, Connors BW (2007). Neuroscience: Exploring the Brain. Lippincott Williams & Wilkins. p. 718. ISBN 0-7817-6003-8.
- ↑ 36,0 36,1 Cooke SF, Bliss TV (2006). "Plasticity in the human central nervous system". Brain. 129 (Pt 7): 1659–73. doi:10.1093/brain/awl082. PMID 16672292.
- ↑ Bliss TV, Collingridge GL (Januarie 1993). "A synaptic model of memory: long-term potentiation in the hippocampus". Nature. 361 (6407): 31–9. doi:10.1038/361031a0. PMID 8421494.
- ↑ Kauer JA, Malenka RC (November 2007). "Synaptic plasticity and addiction". Nat. Rev. Neurosci. 8 (11): 844–58. doi:10.1038/nrn2234. PMID 17948030.
- ↑ 39,0 39,1 Sherrington CS (1906). The Integrative Action of the Nervous System. Scribner.
- ↑ Descartes R (1989). Passions of the Soul. Voss S. Hackett. ISBN 978-0-87220-035-7.
- ↑ Baum WM (2005). Understanding behaviorism: Behavior, Culture and Evolution. Blackwell. ISBN 978-1-4051-1262-8.
- ↑ Piccolino M (November 2002). "Fifty years of the Hodgkin-Huxley era". Trends Neurosci. 25 (11): 552–3. doi:10.1016/S0166-2236(02)02276-2. PMID 12392928.
- ↑ Johnston D, Wu SM (1995). Foundations of cellular neurophysiology. MIT Press. ISBN 978-0-262-10053-3.
- ↑ Dayan P, Abbott LF (2005). Theoretical Neuroscience: Computational and Mathematical Modeling of Neural Systems. MIT Press. ISBN 978-0-262-54185-5.
- ↑ Simmons PJ, Young D (1999). "Ch 1.: Introduction". Nerve cells and animal behaviour. Cambridge Univ. Press. ISBN 978-0-521-62726-9.
- ↑ 46,0 46,1 "Ch. 36: Spinal reflexes". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ "Ch. 38: Voluntary movement". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ "Ch. 39: The control of gaze". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ "Ch. 21: Coding of sensory information". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ "Ch. 25: Constructing the visual image". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ "Ch. 52: The induction and patterning of the nervous system". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ Sanes DH, Reh TH, Harris WA (2006). "Ch. 1, Neural induction". Development of the Nervous System. Elsevier Academic Press. ISBN 978-0-12-618621-5.
- ↑ "Ch. 53: The formation and survival of nerve cells". Principles of Neural Science. McGraw-Hill Professional. 2000. ISBN 978-0-8385-7701-1.
{{cite book}}: Onbekende parameter|editors=geïgnoreer (hulp) - ↑ "ACGME Program Requiremeg" (PDF). Geargiveer vanaf die oorspronklike (PDF) op 13 Januarie 2017. Besoek op 10 Januarie 2017.
Eksterne skakels
[wysig | wysig bron] Wikimedia Commons het meer media in die kategorie Senuweestelsel.
Wikimedia Commons het meer media in die kategorie Senuweestelsel.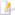 Hierdie artikel is vertaal uit die Engelse Wikipedia
Hierdie artikel is vertaal uit die Engelse Wikipedia