Ranunculales
Ranunculales | ||
|---|---|---|
 | ||
| Clasificación científica | ||
| Reinu: | Plantae | |
| División: | Angiospermae | |
| Clas: | Eudicotyledoneae (basal) | |
| Orde: |
Ranunculales Dumortier | |
| Families | ||
| ||
| Consultes | ||
| World Flora Online | World Flora online | |
| [editar datos en Wikidata] | ||
Ranunculales ye un taxón d'eudicotiledónees allugáu na categoría d'orde, que nos sistemes de clasificación actuales como l'APG (1998),[1] el so socesor APG II (2003),[2] y el más actualizáu APW[3] ta compuestu por siete families que les sos rellaciones filoxenétiques conócense bien dende va un tiempu. Esti clado ye una caña basal de les eudicotiledónees y contién cerca d'un 1,6 % de la diversidá de eudicotiledónees.
Calteres
[editar | editar la fonte]Los miembros d'esti clado comparten los siguientes calteres: en dellos miembros la presencia d'O-metilflavonoles, presencia de flavonoles; na corte tresversal del tarmu nuevu ("cercu") vense los fexes vasculares separaos, los vasos presentes namái na parte central, presencia de traqueides verdaderes; los rayos puramente anchamente multiseriados (y tamién nel tochu, cuando esiste); madera fluorescente en dellos representantes; cámbium estratificado; les célules cribosas del floema con plástidos largamente de tipu S (S-type), presencia de proteína P dispersiva (dispersive P-protein); na corte tresversal ("cercu") del peciolu los fexes vasculares disponer en forma d'aníu; les ceres de la cutícula de la fueya formando grupos de túbulos, siendo la cera principal el nonacosan-10-ol; fueyes dispuestes n'espiral; les pieces del xinecéu opuestes a les del perianto; el perianto deciduo cuando se forma'l frutu; grana con exotesta.
La berberina, de presencia común nesti orde, ye sintetizada pela vía de la tirosina.
A la fecha d'edición d'esti artículu (abril del 2007), nun había información abonda sobre los elementos de vasu, los dientes de les fueyes, el desenvolvimientu del endosperma, nin el tamañu del embrión.
Diversidá
[editar | editar la fonte]Ranunculales ta conformáu polos siguiente grupos monofiléticos:
Eupteleaceae
[editar | editar la fonte]
Compuesta por un únicu xéneru y dos especies d'arbustos o arbolinos dacuando cultivaos como ornamentales, distribuyir pol este y sur d'Asia. Aisllada del restu de les families de dicotiledónees nos sos calteres, pue ser reconocida poles sos fueyes dispuestes n'espiral, dentaes, con venes grueses, ascendentes y pinadas que van escontra los dientes; les bases del peciolu escavaes, y les flores pequeñes que nun tienen perianto y tienen anteres con un conectivo llargu y carpelos separaos y estipitados. El frutu ye una sámara pequeña con forma de discu, debaxo d'un llau reparar los restos del estigma con forma de cepiyu.
Papaveraceae
[editar | editar la fonte]Papaveraceae son usualmente yerbes con dalgún tipu de exudado, con fueyes carnoses con bases amplies. Les flores son dímeras, y la mota y la corola son claramente estremables, el xinecéu tien dos (o más) carpelos, placentación parietal, y usualmente un estilu curtiu.
- Pteridophylloideae son yerbes con una roseta de fueyes bien similares a frondes de felechu, y una inflorescencia de tipu escapo con flores bastante pequeñes y dímeras, con un estilu estremable.
- Papaveroideae usualmente son yerbes que pueden ser reconocíes poles sos fueyes usualmente nidies (bristly-hairy leaves), látex bayurosu, y flores con dos sépalos grandes y fugaces que zarren el brotu, pétalos engurriaos, usualmente numberosos estambres, y un xinecéu sincarpo con como muncho un estilu curtiu y placentación parietal.
- Fumarioideae son usualmente yerbes carnoses con cazumbre enaguada y unes flores distintives dímeras monu o asimétriques que usualmente tán spurred, la mota ye bien pequeñu o casi invisible.
Circaeasteraceae
[editar | editar la fonte]Circaeasteraceae ye fácilmente reconocible. Son yerbes pequeñes, con fueyes con venación largamente dicotómica, y les sos fueyes bastante pequeñes tienen carpelos separaos.
Lardizabalaceae
[editar | editar la fonte]
Lardizabalaceae son usualmente lianas con fueyes compuestes y madera con rayos amplios, les plantes son monoicas o dioicas. Les flores son de tamañu mediu y trímeras, los miembros del perianto opuestos al androcéu, los carpelos son llibres y los frutos son más o menos carnosos, dacuando son folículos.
Menispermaceae
[editar | editar la fonte]
Menispermaceae ye reconocida pol so vezu usualmente de tipo viña o liana, rayos amplios, pecíolos pulvinados nos dos puntes, y usualmente la llámina coriácea con venación palmetada. La llámina tamién ye usualmente peltada, y entá si nun lu fora, los dos llaos de la llámina casi s'atopen na punta del peciolu. Les plantes son dioicas, les flores usualmente trímeras son pequeñes, y el frutu son "drupelets" que usualmente tienen un endocarpo esculpíu de forma distintiva, y puede tar fuertemente curvado (por eso conocer como "la familia de la grana con forma de lluna", cuantimás por cuenta de Menispermum canadense).
Berberidaceae
[editar | editar la fonte]
Berberidaceae son yerbes o arbustos que pueden ser reconocíos pol so texíu secundariu usualmente mariellu (ye más obviu nos raigaños), y les sos fueyes usualmente compuestes y con estípules, con venación palmetada y márxenes fuertemente dentaos o espinosos. Les sos flores son usualmente trímeras, el perianto ye usualmente multiseriado, los estambres opuestos al perianto, y la dehiscencia de l'antera ye valvada, siempres hai un solu carpelo.
Ranunculaceae
[editar | editar la fonte]
Ranunculaceae puede ser reconocida pol so vezu usualmente herbal y les sos fueyes n'espiral, usualmente ensin estípules, llobaes, con bases amplies. Les flores tienen sépalos los grandes y usualmente petaloides, "pétalos" nectaríferos, n'espiral, usualmente numberosos estambres, y carpelos separaos nacíos nun receptáculu bien desenvueltu.
Filoxenia
[editar | editar la fonte]Ranunculales ye'l clado basal de eudicotiledónees. Se hipotetiza que ye monofilético sobre la base de les secuencies d'ADN rbcL, atpB y ADNr 18S (Chase et al. 1993,[4] Drinnan et al. 1994,[5] Hoot y Crane 1995,[6] Loconte et al. 1995,[7] Soltis et al 1997,[8] 1998,[9] 2000,[10] Källersjö et al. 1998,[11] Hoot et al. 1999,[12] Savolainen et al 2000a,[13] b).[14] La presencia del alcaloide berberina tamién puede ser una sinapomorfía del grupu. L'orde ta compuestu por siete clados a los que-yos foi asignada la categoría de familia, les families más numberoses son Menispermaceae, Berberidaceae, Ranunculaceae y Papaveraceae. Estes families tradicionalmente fueron acomuñaes pol so vezu predominantemente herbal, les sos fueyes dentaes o llobaes o hasta compuestes, la presencia d'alcaloides, típicamente del tipu de la bencil-isoquinolina, les flores hipóginas coles partes usualmente estremaes y llibres, y usualmente con munchos estambres, y granes con embriones pequeños y endosperma bayurosu (Cronquist 1981,[15] Thorne 1974,[16] 1992).[17]
El grupu foi usualmente acomuñáu col complexu de magnólidas (por casu coles Magnoliales, les Laurales y les Canellales: Cronquist 1968,[18] 1981,[15] 1988,[19] Dahlgren 1983,[20]Takhtajan 1969,[21] 1980,[22] Thorne 1974,[16] 1992),[17] pero la morfoloxía (Donoghue y Doyle 1989[23] Doyle et al. 1994)[24] y les secuencies d'ADN rbcL, atpB y ADNr 18S yá citaes indiquen que Ranunculales ye'l clado hermanu de tol restu de les tricolpadas (eudicotiledónees).
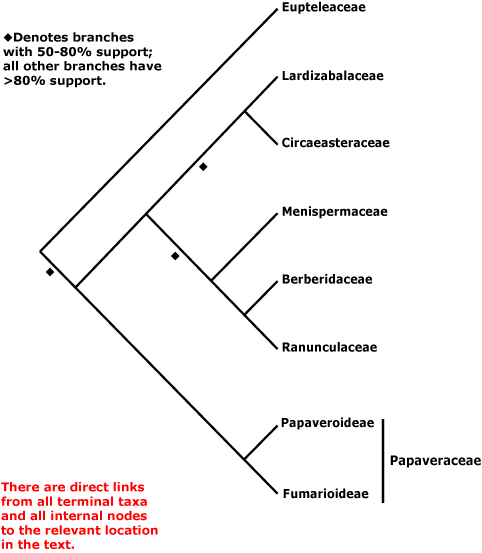
L'árbol filoxenéticu actualizáu hasta abril de 2007:[25]
|
La páxina Plantía:Clade/styles.css nun tien conteníu.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Papaveraceae ta formada por Papaveroideae, Fumarioideae y Pteridophilloideae. Difier del restu de los miembros del grupu nel so xinecéu sincarpo, frutos capsulares, sépalos los tempranamente deciduos, y presencia d'o bien laticíferos y cazumbre coloriada, o bien célules especializaes con cazumbre mucilaginosa y clara. Estos calteres, xunto coles granes con arilo, son probablemente sinapomorfías de Papaveraceae, o evolucionaron bien tempranamente nesta familia.
Les sinapomorfías morfolóxiques del clado qu'axunta a Menispermaceae, Berberidaceae y Ranunculaceae nun son evidentes, anque'l grupu tien un robezu soporte per parte de les secuencies d'ADN. Pue que Berberidaceae y Ranunculaceae tengan delles semeyances na forma de la flor (Endress 1995).[26]
Taxonomía
[editar | editar la fonte]- Introducción teórica en Taxonomía
L'orde contién 7 families, 199 xéneros, y 4445 species.
- Familia Ranunculaceae
- Familia Berberidaceae
- Familia Menispermaceae
- Familia Lardizabalaceae
- Familia Circaeasteraceae
- Familia Eupteleaceae
- Familia Papaveraceae (incluyendo a Fumariaceae y Pteridophyllaceae)
Esisten otres poques families separaes del orde Ranunculaceae. Nel sistema de clasificación de Cronquist (Cronquist 1981,[15] 1988),[27] les Eupteleaceae yeren escluyíes d'esti orde, como tamién les Papaveraceae y Fumariaceae (incluyendo a les Pteridophyllaceae) que yeren consideraes como parte del orde Papaverales.
Sinónimos:
- Berberidales Dumortier, *
Eupteleales Reveal, * Glaucidiales Reveal, * Helleborales Nakai, * Hydrastidales Takhtajan, * Lardizabalales Loconte, * Menispermales Bromhead, * Nandinales Doweld, * Papaverales Dumortier, * Podophyllales Dumortier - Berberidanae Doweld, * Ranunculanae Reveal - Ranunculidae Reveal - Berberidopsida Brogniart, * Papaveropsida Brongniart, * Ranunculopsida Brongniart
Evolución
[editar | editar la fonte]Magallön et al. (1999)[28] suxeren que'l clado tien una edá de 70 Ma, basaos nos fósiles más antiguos que pueden ser asignaos al mesmu, pero esos fósiles son miembros de Menispermaceae que nun ye un lineaje basal. Anderson et al. (2005)[29] daten al ancestru de Ranunculales y tolos sos parientes próximos yá estinguíos (el "stem group" Ranunculales) nunos 122-120 Ma, y la diverxencia yá dientro del clado nunos 121-114 Ma. Toles families diverxeron antes de los 105 Ma sacante la división ente Ranunculaceae y Berberidaceae qu'asocedió hai unos 104-90 Ma (ver Anderson et al. 2005 pa más detalles de diversificaciones posteriores dientro del clado).
Gleissberg y Kadereit (1999)[30] alderiquen la evolución de la morfoloxía de les fueyes nel clado, y consideren que les fueyes de tipu ancestral (plesiomórficas) quiciabes fueren politernada, acrópeta, y basípetamente pedada. Los dientes de les fueyes glandulares, con un claru, persistente, "swollen cap" (una especie de capuchón que tien el diente, y que frecuentemente lo fai paecer un botoncito redondu, y suel esconder un hidatodo) al cual tamién lleguen les venes llaterales d'ordes más altos. Con al respective de los coléteres, Drinnan et al. (1994)[31] suxeren que l'apaición de pétalos a partir d'estambres tien d'asoceder munches vegaes dientro del clado, por casu en Lardizabalaceae. Tamura (1965)[32] resume la evidencia pola naturaleza estaminodial de los pétalos en Ranunculaceae: los pétalos tienen un solu trazu foliar, tán nos mesmos parásticos que los miembros del androcéu, y son similares a los estambres nel so desenvolvimientu tempranu, y tamién usualmente son peltados (con pie). Con al respective de si la endexina del polen ye lamelada, el APW remarca que les célules de les antípodes de normal nun son a cencielles persistentes; puede atopase una llista de datos en Williams y Friedman (2004).[33]
Ecoloxía
[editar | editar la fonte]Esti clado, quiciabes especialmente Menispermaceae y Ranunculaceae, nun ye bien usáu poles caparines p'alimentación de les poliyes (Ehrlich y Raven 1964[34]). Hai datos alrodiu de una fertilización retrasada por dos meses o más dempués d'asocedida la polinización, en clados como Eupteleaceae, Circeasteraceae, Lardizabalaceae, y Ranunculaceae (ver Sogo y Tobe 2006[35] y referencies).
Referencies
[editar | editar la fonte]- ↑ Angiosperm Phylogeny Group. 1998. An ordinal classification for the families of flowering plants. Ann Misouri Bot. Gard. 85: 531-553.
- ↑ The Angiosperm Phylogeny Group. 2003. «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II.» Botanical Journal of the Linnean Society, 141, 399-436. (pdf equí (enllaz rotu disponible n'Internet Archive; ver l'historial y la última versión).)
- ↑ Stevens, P. F. 2001 Angiosperm Phylogeny Website, versión 7 (mayu de 2006) y actualizáu regularmente dende entós.
- ↑ Chase, M. W. Soltis, D. E., Olmstead, R. G., Morgan, D., les, D. H., Mishler, B. D., Duvall, M. R., Price, R. A., Hills, H. G., Qiu, Y.-L., Kron, K. A., Rettig, J. H., Conti, E., Palmer, J. D., Manhart, J. R., Sytsma, K. J., Michaels, H. J., Kress, W. J., Karol, K. G., Clark, W. D., Hedrén, M., Gaut, B. S., Jansen, R. K., Kim, K.-J., Wimpee, C. F., Smith, J. F., Furnier, G. R., Strauss, S. H., Xiang, Q.-Y., Plunkett, G. M., Soltis, P. S., Swensen, S. M., Williams, S. E., Gadek, P. A., Quinn, C. J., Eguiarte, L. E., Golenberg, E., Learn, G. H., Jr., Graham, S. W., Barrett, S. C. H., Dayanandan, S., y Albert, V. A. 1993. «Phylogenetics of seed plants: An analysis of nucleotide sequences from the plastid gene rbcL.» Ann. Missouri Bot. Gard. 80: 528-580.
- ↑ Drinnan, AN, PR Crane, y SB Hoot. 1994. P«atterns of floral evolution in the early diversification of non-magnoliid dicotyledons (eudicots).» Plant Syst. Evol. Suppl 8:93-122.
- ↑ Hoot, SB y PR Crane. 1995. «Interfamilial relationships in the Ranunculidae based on molecular systematics.» Plant Syst. Evol. Suppl. 9: 119-131.
- ↑ Loconte H, LM Campbell, y DW Stevenson. 1995. «Ordinal and familial relationships of ranunculid genera.» Plant Syst Evol. Suppl. 9: 99-118.
- ↑ Soltis, D. E. Kuzoff, R. K., Nickrent, D. L., Johnson, L. A., Hahn, W. J., Hoot, S. B., Sweere, J. A., Kuzoff, R. K., Kron, K. A., Chase, M. W., Swensen, S. M., Zimmer, E. A., Chaw, S.-M., Gillespie, L. J., Kresss, W. J., y Sytsma, K. J. 1997.« Angiosperm phylogeny inferred from 18S ribosomal DNA sequences.» Ann. Missouri Bot. Gard. 84: 1-49.
- ↑ Soltis DE, PS Soltis, ME Mort, MW Chase, V Savolainen, SB Hoot y CM Morton. 1998. «Inferring complex phylogenies using parsimony: An empirical approach using three large DNA data sets for angiosperms.» Syst Biol. 47: 32-42.
- ↑ Soltis DE, PS Soltis, MW Chase, ME Mort, DC Albach, M Zanis, V Savolainen, WH Hahn, SB Hoot, MF Fay, M Axtell, SM Swensen, LM Prince, WJ Kress, KC Nixon, y JS Farris. 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Bot. J. Linn. Soc. 133: 381-461. (resume equí)
- ↑ Källersjö M, JS Farris, MW Chase, B Bremer, MF Fay,,CJ Humphries, G Petersen, O Seberg, y K Bremer. 1998. «Simultaneous parsimony jacknife analysis of 2538 rbcL DNA sequences reveals support for major clades of green plants, land plants, and flowering plants.» Pl. Syst. Evol. 213: 259-287.
- ↑ Hoot SB, S Magallón, y PR Crane. 1999. «Phylogeny of basal eudicots based on three molecular data sets: atpB, rbcL, and 18S nuclear ribosomal DNA sequences.» Ann. Missouri Bot. Gard. 86: 1-32.
- ↑ Savolainen V, MW Chase, SB Hoot, CM Morton, DE Soltis, CBayer, MF Fay, AI de Brujin, S Sullivan, y Y-l Qiu. 2000a. «Phylogenetics of flowering plants based upon a combined analysis of plastid atpB and rbcL gene sequences.» Syst Biol. 49: 306-362.
- ↑ Savolainen V, MF Fay, DC Alback, A Backlund, M van der Bank, KM Cameron, SA Johnson, MD Lledó, J-C Pintaud, M. Powell, MC Sheahan, DE Soltis, PS Soltis, P Weston, WM Whitten, KJ Wurdack, y MW Chase. 2000b. «Phylogeny of the eudicots: a nearly complete familial analysis based on rbcL gene sequences.» Kew Bull. 55: 257-309.
- ↑ 15,0 15,1 15,2 Cronquist, A. 1981. An integrated system of classification of flowering plants. Columbia University Press, Nueva York.
- ↑ 16,0 16,1 Thorne, R. F. 1974. «A phylogenetic classification of the Annoniflorae.» Omeru 8: 147-209.
- ↑ 17,0 17,1 Thorne, R. F. 1992. «Classification and geography of the flowering plants.» Bot Rev. 58: 225-348.
- ↑ Cronquist, A. 1968. The evolution and classification of flowering plants. Houghton Mifflin, Boston.
- ↑ Cronquist, A. 1988. The evolution and classification of flowering plants. 2ª edición. New York Botanical Garden, Bronx.
- ↑ Dahlgren, R. M. T. 1983. Xeneral aspects of angiosperm evolution and macrosystematics. Nordic. J. Bot. 3: 119-149.
- ↑ Takhtajan, A. 1969. Floweringplants: Origin and dispersal. Smithsonian institution press. Washington, DC.
- ↑ Takhtajan, A. 1980. Outline of the classification of floweing plants (Magnoliophyta). Bot. Rev.'}' 46: 225-359.
- ↑ Donogue MJ y JA Doyle. 1989. Phylogeneticc analysis of angiosperms and the relationships of Hamamelidae. En: Evolution, systematics, and fossil history of the Hamamelidae, vol. 1, Introduction and "lower" Hamamelidae. Syst. Assoc. Special Vol. 40A, P. R. Crane y S. Blackmore (eds.), 17-45. Clarendon Press, Oxford.
- ↑ Doyle JA, MJ Donoghue y EA Zimmer. 1994. Integration of morphological and ribosomalRNA data on the origin of angiosperms. Ann. Missouri Bot. Garden81: 419-450.
- ↑ Árbol en APW
- ↑ Endress, PK. 1995. Floral structure and evolution in Ranunculanae. Plant Syst. Evol. Suppl. 9: 47-61.
- ↑ Cronquist, A. 1988. The evolution and classification of flowering plants. 2nd ed. New York Botanical Garden, Bronx.
- ↑ Magallón, S. A., Crane, P. R., y Herendeen, P. S. 1999. Phylogenetic pattern, diversity, and diversification of eudicots. Ann. Missouri Bot. Gard. 86: 297-372.
- ↑ Anderson, C. L., Bremer, K., y Friis, Y. M. 2005. Dating phylogenetically basal eudicots using rbcL sequences and multiple fossil reference points. American J. Bot. 92: 1737-1748.
- ↑ Gleissberg, S., y Kadereit, J. W. 1999. Evolution of leaf morphogenesis: Evidence from developmental and phylogenetic data in Papaveraceae. Int. J. Plant Sci. 160: 787-794.
- ↑ Drinnan, Crane, y Hoot, S. B. 1994. Patterns of floral evolution in the early diversification of non-magnoliid dicotyledons (eudicots). Pp. 93-122, en: Endress, P. K., y Friis, Y. M. (eds.), Early Evolution of Flowers. Springer, Vienna. [Plant Syst. Evol. Suppl. 8.]
- ↑ Tamura, M. 1965. Morphology, ecology and phylogeny of the Ranunculaceae IV. (Ranunculaceae of Eastern Asia: Xeneral Part IV). Sci. Rep. Osaka Univ. 14(1): 53-71.
- ↑ Williams, J. H., Jr., y Friedman, W. Y. 2004. The four-celled female gametophyte of Illicium (Illiciaceae; Austrobaileyales): Implications for understanding the origin and early evolution of monocotyledons, eumagnoliids, and eudicots. American J. Bot. 91: 332-351.
- ↑ Ehrlich, P. R., y Raven, P. H. 1964. Butterflies and plants: A study in coevolution. Evolution 18: 586-608.
- ↑ Sogo, A., y Tobe, H. 2006. Delayed fertilization and pollen-tube growth in pistils of Fagus japonica (Fagaceae). American J. Bot. 93: 1748-1756.
{kind=link}
Bibliografía
[editar | editar la fonte]- Ranunculales en: Stevens, P. F. 2001 Angiosperm Phylogeny Website, versión 7 (mayu de 2006) y actualizáu regularmente dende entós. Fecha d'accesu: 29 d'abril de 2007.
- "Ranunculales". en: Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA. Capítulu 9, "Phylogenetic Relationships of Angiosperms".
Enllaces esternos
[editar | editar la fonte]| Wikispecies tien un artículu sobre Ranunculales. |
 Wikimedia Commons acueye conteníu multimedia sobre Ranunculales.
Wikimedia Commons acueye conteníu multimedia sobre Ranunculales.
| Control d'autoridaes |
|
|---|
 Datos: Q21021
Datos: Q21021- Multimedia: Ranunculales
 Especies: Ranunculales
Especies: Ranunculales