Lectin




Lectiner er kulhydrat-bindende proteiner med vigtige funktioner i mennesker, dyr, planter og mikroorganismer.[1][2][3][4] Lectiner benævnes også agglutininer pga. deres evne til at agglutinere celler, som f.eks. hæmagglutininer eller fytohæmagglutininer. Lectiner renfremstilles fra planter, svampe og enkelte andre organismer og anvendes i medicinen og biokemien til analyser og processer involverende kulhydrater og kulhydrat-indeholdende molekyler. Mange lectiner er naturligt forekommende toksiner.[5]
Lectinet ricin fra castorbønnen blev beskrevet som det første lectin af den estiske biokemiker Peter Hermann Stillmark i hans disputats fra 1888: "Ueber Ricin, ein giftiges Ferment aus den Samen von Ricinus comm. L. und einigen anderen Euphorbiaceen". Beskrivelsen blev starten til en ny gren af naturvidenskaben kaldet lectinologi.
På grund af lectinernes biologiske aktivitet som pesticider, er fødemidler blevet genetisk modificeret med lectin-gener. Således indeholder vintergækker et aktivt lectin eller agglutinin benævnt GNA (for Galanthus nivalis agglutinin). Der er bl.a. udført forsøg med GNA-genet overført til kartofler. Herom sagde i 1998 Árpád Pusztai, en skotsk-ungarsk biokemiker og ernæringsforsker, i et interview på britisk TV (World in Action), at hans forskningsgruppe havde observeret skader på tarmsystemet og immunsystemet på rotter fodret med de genetisk modificerede kartofler. Bl.a. sagde han: "If I had the choice I would certainly not eat it", og "I find it's very unfair to use our fellow citizens as guinea pigs".[6] Disse bemærkninger startedes Pusztai-sagen om genetisk modificerede fødemidler.
Ordforklaring
[redigér | rediger kildetekst]- CRD = carbohydrate recognition domain
- C-type lectin = lectin der kræver Ca2+ for binding
- S-type lectin = lectin der kræver disulfidbroer for stabilitet og binding
Animalske lektiner
[redigér | rediger kildetekst]Mennesket producerer over 200 lectiner, alle med vigtige roller i organismens normale funktion som celle-til-celle adhæsion og apoptose. Lectinerne fungerer også som en arm af immunsystemet, både den medfødte og den adaptive immunitet. Nogle lectiner opdager mikrober - bakterier, virus, svampe og protozoer - og medierer deres direkte tilintetgørelse eller signalerer deres tilstedeværelse til immunsystemet, idet de er lokaliseret hvor organismen møder mikroorganismer bl.a. i huden, i de øvre luftveje, og i den gastrointestinale og den reproduktive kanal. På slimhinderne virker lectinerne sammen med bl.a. muciner, antimikrobielle peptider også kaldet defensiner, immunglobuliner og andre faktorer for at forhindre mikroorganismer i at gennemtrænge epitelbarrieren. En fyldig oversigt over animalske lectiner findes her.[7]
Collectiner
[redigér | rediger kildetekst]Collectiner er lectiner af C-type, der delvist består af et kollagent domæne, der kan danne en tredobbelt helix som basis for trivalente og multivalente strukturer med binding til mikroorganismer. Bindingen kan udløse eliminering af mikroorganismerne på en række måder: aggregering, aktivering af komplement, opsonisering, fagocytose eller hæmning af den mikrobielle vækst. Andre af collectinernes funktioner er modulering af inflammatoriske, allergiske reaktioner og clearing af apoptotiske celler.
Ni typer collectiner er defineret:
- MBL = MBP, det mannose-bindende lectin (se nedenfor)
- SP-A = surfactant protein A
- SP-D = surfactant protein D [8]
- CL-L1 = collectin liver 1
- CL-P1 = collectin placenta 1
- CL-43 = collectin of 43 kDa
- CL-46 = collectin of 46 kDa
- CL-K1 = collectin kidney 1
- Conglutinin
MBP, det mannose-bindende lectin
[redigér | rediger kildetekst]MBP eller MBL (det mannose-bindende lectin eller det mannan-bindende lektin) er et naturligt forekommende plasma protein, som spiller en hovedrolle i det første immunforsvar mod infektioner, i Komplementsystemets lectinreaktionsvej. For eksempel beskytter MBL det nyfødte barn i de første dage efter fødslen indtil det normale antistof-baserede immunsystem begynder at virke. MBL virker som organismens ildslukker mens der ventes på brandvæsnet. De fleste mennesker har et normalt indhold i blodet af MBL, men for en ret stor del af befolkningen (op til 40%) er der nedsat indhold af MBL på grund af genetiske mutationer. Det kan være kritisk under sygdomsforhold, f.eks. for kræftpatienter under kemoterapi og for cystisk fibrose patienter.
Galectiner
[redigér | rediger kildetekst]Galectiner, tidligere kaldt S-type-lektiner, er en klasse af lektiner, der binder specifikt til β-galactosid-forbindelser, såsom N-acetyllactosamin (Galβ1-3GlcNAc eller Galβ1-4GlcNAc), som findes vidt udbredt blandt animale proteiner ved enten N-bundet eller O-bundet glycosylering. Galectinerne har en bred vifte af funktioner, herunder apoptose, celle-celle-interaktioner, celle-matrix-adhæsion og transmembran-signalering.
Der er blevet opdaget omkring 15 galectiner i pattedyr, kodet af LGALS-generne, som er nummereret på en fortløbende måde. Kun galectin-1, -2, -3, -4, -7, -7B, -8, -9, -9B, 9C, -10, -12, -13, -14 og -16 er blevet identificeret i mennesker. Medlemmer af galectinfamilien er repræsenteret i pattedyr, fugle, padder, fisk, nematoder og nogle svampe.[9]
Ficoliner
[redigér | rediger kildetekst]Ligesom collectinerne er ficolinerne trimere proteiner med kollagen-lignende strukturer, der binder mikrober og aktiverer lectin-vejen for komplement. Ficolinerne reagerer med kulhydrater ved hjælp af et fibrinogen-lignende domæne. Den primære ficolinstruktur indeholder 288 aminosyrer. Kombinationen af kollagen-lignende og fibrinogen-lignende domæne tillader proteinet at danne en grundlæggende underenhed indeholdende en tredobbelt spiralformet hale og en trio af kugleformede hoveder. Ficoliner produceres i leveren af hepatocytter og i lungerne af alveolære celler type II, neutrofiler og monocytter.
Sammen med pentraxiner, collectiner og C1q-molekyler udgør ficoliner de opløselige lectiner for mønstergenkendelse af mikroorganismernes overflademolekyler.
H-Ficolin eller ficolin3 ser ud til at være det mest udbredte lectin i cirkulationen, med koncentrationer fra 20 til 30 μg/ml. Det er også til stede i tyktarmen og lungerne. H-Ficolin-mangel resulterer i immundefekt.
Pentraxiner
[redigér | rediger kildetekst]Pentraxiner er lektiner karakteriseret ved en cyklisk pentamer struktur fundet i dyr fra insekter til mennesker og er klassificeret som “korte” (CRP eller C-reactive protein og SAP eller Serum amyloid P component) eller “lange” (PTX3). Ligesom collectinerne kan pentraxiner rekruttere komplement og lette antigenoptagelsen. CRP var et af de første humorale lectiner, der blev identificeret. Den normale serumkoncentration på 3 μg/ml kan stige 1.000 gange ved infektion, hvorfor CRP bruges som markør for betændelse og infektion. Dette lectin dannes også i lungerne.
Asialoglycoprotein-receptoren
[redigér | rediger kildetekst]Asialoglycoprotein-receptoren er et klassisk eksempel på den fysiologiske betydning af N-glycaner. Denne receptor er et membranprotein på overfladen af levercellerne binder glycoproteiner, der har mistet den normale terminale sialinsyre, dvs. binder eksponerede galactosylrester og endocyterer dem og viser den fysiologiske clearings-mekanisme af serumglycoproteiner.[10]
Andre animalske lectiner
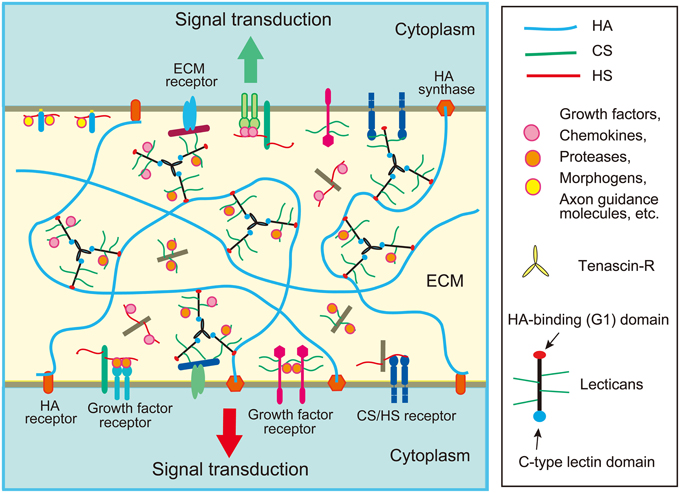
[redigér | rediger kildetekst]- Aggrecan
- Brevican
- CLR, C-Type lectin receptors[11]
- Intelectiner[12]
- LBP, lipopolysaccharide-binding protein[13]
- Lecticaner[14]
- Neurocan[15]
- Reg, regenerating islet-derived protein[16]
- Selectiner[17]
- Tetranectin[18]
- TFF, trefoil factor[19]
- Versican
- ZG, zymogen granule.
Phytohæmagglutininer, plante-lectiner
[redigér | rediger kildetekst]| Tabel over visse plante- og svampelectiner[20] | |||||
|---|---|---|---|---|---|
| Symbol | Lectin-navn | Kilde | Ligand, for flere detaljer se [20] | ||
| Mannose-bindende lectiner | |||||
| ConA | Concanavalin A, se en:Concanavalin A | Canavalia ensiformes, springbønne | α-D-mannose og α-D-glucose | ||
| LCH | Lentil lectin | Lens culinaris | bi- og triantenne N-glycaner | ||
| GNA | "Snowdrop lectin", vintergæk-lectin | Galanthus nivalis | α 1-3 og α 1-6 linked high mannose structurer | ||
| Galaktose / N-acetylgalaktosamin-bindende lectins | |||||
| RCA | Ricin, se en:Ricin, Ricinus communis Agglutinin, RCA120 | Ricinus communis | Galβ1-4GlcNAcβ1-R | ||
| PNA | "Peanut agglutinin", jordnød-lectin, se en:Pesnut agglutinin | Arachis hypogaea | Galβ1-3GalNAcα1-Ser/Thr (T-Antigen) | ||
| AIL | Jacalin, se en:Jacalin | Artocarpus integrifolia | (Sia)Galβ1-3GalNAcα1-Ser/Thr (T-Antigen) | ||
| VVL | "Hairy vetch lectin" | Vicia villosa, Sand-vikke | GalNAcα-Ser/Thr (Tn-Antigen) | ||
| N-acetylglukosamin-bindende lectiner | |||||
| WGA | "Wheat Germ agglutinin", hvedkim-lectin, se en:WGA | Triticum vulgaris | GlcNAcβ1-4GlcNAcβ1-4GlcNAc, Neu5Ac (sialic acid) | ||
| N-acetylneuraminsyre-bindende lectiner | |||||
| SNA | "Elderberry lectin", hyld-lectin | Sambucus nigra | Neu5Acα2-6Gal(NAc)-R | ||
| MAL | Maackia amurensis leukoagglutinin | Maackia amurensis | Neu5Ac/Gcα2,3Galβ1,4Glc(NAc) | ||
| MAH | Maackia amurensis hemoagglutinin | Maackia amurensis | Neu5Ac/Gcα2,3Galβ1,3(Neu5Acα2,6)GalNac | ||
| Fukosebindende lectiner | |||||
| UEA | Ulex europaeus agglutinin | Ulex europaeus | Fucα1-2Gal-R | ||
| AAL | Aleuria aurantia lectin | Aleuria aurantia | Fucα1-2Galβ1-4(Fucα1-3/4)Galβ1-4GlcNAc, R2-GlcNAcβ1-4(Fucα1-6)GlcNAc-R1 | ||


De først kendte lectiner var fra planter og blev kaldt phytohæmagglutininer pga. deres evne til at agglutinere røde blodlegemer. Der kendes nu et utal af lectiner fra planter, nogle få er nævnt i tabellen.
Lectin som insekticid
[redigér | rediger kildetekst]Bacillus thuringiensis forkortet til Bt er en almindelig spore-dannende Gram-positiv jordbakterie, der under sporuleringen danner protein-krystaller af et eller flere lectiner, der benævnes Cry-toksiner (Cry for krystal) eller δ-endotoksiner. Cry-toksinerne kodes af plasmidgenerne Cry og er toksiske for insekter, specielt larver af Lepidoptera (sommerfugle) samt Coleoptera (biller) og Diptera (tovinger (fluer)). Den toksiske virkning ytrer sig gennem en nedbrydning af tarmen.[21] Toksinerne har en fælles struktur af tre domæner hvor domæne II og domæne III har lectin-lignende aktivitet, der bestemmer bindingen til de modtagelige insekters tarm.[22]
Da Bt-toksinerne ikke anses for at være skadelige for mennesker[23] er afgrøder som majs, bomuld og kartofler blevet genetisk modificeret med Bt-toksiner.[24] Da Bt-toksinerne har lille effekt på andre organismer, er brugen af Bt-transgene afgrøder blevet anset for at være mere miljøvenlig end brugen af syntetiske pesticider, men indførelsen og brugen af de gensplejsede afgrøder har affødt en stående diskussion jf. genetisk modificeret organisme.[25]
Andre giftige lectiner
[redigér | rediger kildetekst]Igennem mange år har det været diskuteret, om lectiner i madvarer ud over deres toxiske virkning også er sygdomsfremkaldende faktorer.[26][27]
Alle bønnespirer og bønner bortset fra linser, mungbønner, soyabønner og kikærter er giftige i rå tilstand på grund af lectinindholdet. Lectinerne i havebønner, pralbønner, limabønner, hestebønner, hvide bønner, brune bønner, sorte bønner, pintobønner og især kidneybønner giver diarré, opkastning og kvalme, i værre fald mavekrampe og blødninger.[28][29][30]
Alle dele af påskeliljen indeholder et giftigt lectin, men særligt løget har et højt indhold.[31] Løgplanter indeholder alle formentlig giftige lectiner i lighed med påskelilje og vintergæk.
Pokeweed mitogen, PMW fra Amerikansk Kermesbær agglutinerer som mange andre plantelectiner erytrocytter, og er - som navnet antyder - et mitogen med stimulerende aktivitet og fremmer antistofsyntesen i lymfocytter, inducerer plasmacellers aktivering og kan i stor dosis være dødelig.
Se også
[redigér | rediger kildetekst]- Ebolavirus
- GMO, genmodificerede organismer
- Glycobiologi
- Mitogen
Eksterne links
[redigér | rediger kildetekst]- Proteopedia viser mere end 800 tredimensionale molekylmodeller af lectiner, brudstykker af lectiner og komplekser med kulhydrater
- EY Laboratories, Inc World's largest lectin manufacturer.
- MBL-deficiens. Statens Serum Institut
Referencer
[redigér | rediger kildetekst]- ^ Lectins. Glycoforum
- ^ A glycobiology review: carbohydrates, lectins, and implications in cancer therapeutics. Haike Ghazarian 2011
- ^ The Next Gluten. JAMES HAMBLIN 2017
- ^ The immunomodulatory effect of plant lectins. Maria A. Sousa 2013
- ^ Taylor, Maureen E. (2017). in Essentials of Glycobiology (internet) 3rd edition. Cold Spring Harbor Press. s. Ch. 28.
- ^ "Árpád Pusztai: Biological Divide – James Randerson interviews biologist Árpád Pusztai". London: The Guardian. 15. januar 2008. Hentet 25. april 2010.
- ^ Soluble Human Lectins at the Host–Microbe Interface. ANNUAL REVIEW OF BIOCHEMISTRY 2024
- ^ An Insight into the Diverse Roles of Surfactant Proteins, SP-A and SP-D in Innate and Adaptive Immunity. Frontiers in Immunology 2012
- ^ Galectin functions in cancer-associated inflammation and thrombosis. Frontiers in Cardiovascular Medicine 2023
- ^ [https://pubs.acs.org/doi/pdf/10.1021/bi00495a001 The Asialoglycoprotein Receptor: A Model for Endocytic Transport Receptors. Biochemistry 1990
- ^ C-Type Lectin-Like Receptors: Head or Tail in Cell Death Immunity. Frontiers in Immunology 2020
- ^ Human intelectin-1 (ITLN1) genetic variation and intestinal expression. ScientificReports natureresearch 2021
- ^ Effects of Lipopolysaccharide-Binding Protein (LBP) Single Nucleotide Polymorphism (SNP) in Infections, Inflammatory Diseases, Metabolic Disorders and Cancers. Frontiers in immunology 2021
- ^ Proteoglycans and neuronal migration in the cerebral cortex during development and disease. Frontiers in Neuroscience 2015
- ^ Role of the Chondroitin Sulfate Proteoglycan, Neurocan, in Inhibition of Sensory Neurite Regeneration. UKnowledge 2016
- ^ Four Decades After the Discovery of Regenerating Islet-Derived (Reg) Proteins: Current Understanding and Challenges. Frontiers in Cell Development Biology 2019
- ^ Selectin-mediated Cell Recognition and its Structural Basis. Glycoforum 2002
- ^ ScienceDirect 2014
- ^ Trefoil Factor. ScienceDirect 2014
- ^ a b "Lectin list" (PDF). Interchim. 2010. Hentet 2010-05-05.
- ^ Characterization of Cry toxin mode of action. University of Tennessee, Institute of Agriculture
- ^ Basic Structural Features of the Bacillus thuringiensis crystal toxins
- ^ Glare, T. R., and O´Callaghan, M. 2000. “Bacillus thuringiensis: Biology, Ecology and Safety”. John Wiley and Sons Ltd, Chichester, U.K
- ^ Genetically improved potatoes: protection from damage by Colorado potato beetles. Plant Molecular Biology (Webside ikke længere tilgængelig)
- ^ EU-stormagter nedtromler dansk GMO-forslag. DR 09. mar. 2012
- ^ Do dietary lectins cause disease? David L.J.Freed
- ^ The Lectin Report. Krispin Sullivan
- ^ "Bønner. Camilla Plum: I haven. DR". Arkiveret fra originalen 18. juni 2012. Hentet 7. juli 2012.
- ^ "Proteinrige bønner og linser. Sundhed. DR". Arkiveret fra originalen 23. februar 2014. Hentet 7. juli 2012.
- ^ Bønner er sundt, men husk lige at koge dem først. Dr.dk 2013
- ^ Påskeliljen kan mere end se dejlig ud. Ingeniøren, 2000
{kind=link}