Caudoviricetes (von lateinischcauda‚Schwanz‘) ist eine Klasse von Viren, die Bakterien und Archaeen als Wirte nutzen und eine Kopf-Schwanz-Struktur besitzen, weshalb die Mitglieder der Caudoviricetes einerseits als Bakteriophagen klassifiziert, andererseits gelegentlich informell als Schwanzviren bezeichnet werden.
Die Klasse umfasste bis zum März 2022 nur eine einzige vom International Committee on Taxonomy of Viruses (ICTV) bestätigte Ordnung, Caudovirales.
Diese Ordnung war 1998 von Hans-Wolfgang Ackermann aufgrund von EM-Aufnahmen der Virionen als Supergruppe aller geschwänzten Bakteriophagen vorgeschlagen worden, was in der Folge vom ICTV bestätigt wurde.
Die klassische Unterteilung dieser Gruppe in Familien erfolgte ebenfalls hauptsächlich aufgrund der genauen Morphologie des am Kapsid sitzenden Schwanzstückes:
Myoviren (englischmyoviruses, ehemalige Familie Myoviridae): Morphotyp A: langes kontraktiles Schwanzstück
Siphoviren (en. siphoviruses, ehemalige Familie Siphoviridae): Morphotyp B: langes nicht-kontraktiles Schwanzstück
Podoviren (en. podoviruses, ehemalige Familie Podoviridae): Morphotyp C: kurzes nicht-kontraktiles Schwanzstück.
In zunehmendem Maße erlangen aber Gen- und Genom- bzw. Proteom-Vergleiche für die Taxonomie an Bedeutung.
Es zeigte sich dabei, dass diese drei traditionellen morphologisch begründeten Familien nicht monophyletisch waren. Dies hatte zur Folge, dass Genomsequenzen (sog. Contigs) aus der Metagenomik im bestehenden System – mit fehlender Information über die Morphologie – nicht zugeordnet werden konnten.[6]
Um dieses Problem zu lösen, hatte das ICTV zunächst verschiedene Gruppen aus diesen herkömmlichen Familien – zunächst formell noch innerhalb der damaligen Ordnung Caudovirales – in neue geschaffene Familien verschoben, darunter die Ackermannviridae, Autographiviridae, Chaseviridae, Demerecviridae und Herelleviridae.
Im März 2021 resultierten diese Bestrebungen in dem Vorschlag einer kompletten Reorganisation der Klasse unter Abschaffung der bisherigen Ordnung Caudovirales; sowie der Auflösung der damaligen polphyletischen Familien (Myoviridae, Podoviridae und Siphoviridae), wobei diese durch neue monophyletische Familien ersetzt werden und nur noch informall als nicht-taxonomische Gruppen zur morphologischen Klassifizierung (Morphotypen) bestehen bleiben.[6]
Diesen Vorschlag hat das ICTV dann im März 2022 vollständig umgesetzt. Die Ordnung Caudovirales wurde aufgelöst.
Der Klade der aus der Metagenomik identifizierten crAss-like phages wurde dabei gemäß Vorschlag der Rang einer Ordnung Crassvirales gegeben[6] und noch weitere Ordnungen eingerichtet. Neben den oben genannten bereits aus den herkömmlichen drei Familien (Morphotypen) ausgegliederten Familien wurden dabei weitere Familien definiert, etliche Unterfamilien verblieben aber zunächst noch ohne Familienzuordnung.
Das Genom der Caudoviricetes ist unsegmentiert (monopartit), d. h. es besteht aus einem einzigen Molekül einer (gewöhnlich) doppelsträngigen DNA mit einer Größe von 18 bis 500 kBp (Kilobasenpaare). Es ist bei den klassischen Myo-, Sipho- und Podoviren linear, kann aber (wie bei den Crassvirales, en. crAss-like phages) auch als eine ringförmig geschlossene (zirkuläre) DNA vorliegen.
Neben den üblichen Nukleotiden des genetischen Codes besitzen die Caudoviricetes auch modifizierte, z. B. glykosylierte Basen (d. h. mit angehängten zusätzlichen Zuckerresten) oder 5-Hydroxymethylcytosin an Stelle von Cytosin.
Die DNA befindet sich in einem 45 bis 170 nm im Durchmesser großen ikosaedrischenKapsid aus meist 72 Kapsomeren; das Schwanzstück kann bis zu 230 nm lang sein.
Das Genom kodiert für 27 bis über 600 Gene, deren Anordnung stark variiert. Das Genom kann von einzelsträngigen Lücken durchbrochen sein und im linearen Fall kovalent gebundene terminale Proteine besitzen. Im Bakterium können diese Caudoviricetes in das Nukleoid integrieren (Prophage) oder als lineares bzw. zirkuläres Plasmid in der Zelle verbleiben. Die Anzahl der isolierten Genome ist verhältnismäßig hoch, da die Caudoviricetes einem ständigen genetischen Austausch zwischen viralem und bakteriellem Genom wie auch dem horizontalen Austausch als Plasmid zwischen Bakterien unterliegen (siehe horizontaler Gentransfer). Es entsteht so eine Vielzahl von so genannten „Mosaiktypen“.
Die Caudoviricetes sind stammesgeschichtlich sehr alte Viren mit großen Populationsvarianten und weltweiter Verbreitung. Man schätzt, dass sich etwa 1031Virionen der Caudoviricetes in der Biosphäre befinden, die aneinandergelegt einer Strecke von 2×108Lichtjahren entsprächen. Die Caudoviricetes machen den überwiegenden Anteil des Virioplanktons in den Meeren und Gewässern aus.
Mit Stand Ende April 2023 kennt das International Committee on Taxonomy of Viruses (ICTV) die folgende Familien und Unterfamilien (ergänzt um einige Vorschläge in doppelten Anführungszeichen), wobei etliche Gattungen, Unterfamilien und Familien noch ohne Zuordnung sind:[4]
OrdnungCrassvirales – KladecrAss-like phages mit zirkulärem Genom, ursprünglich als Mitglieder der ehemaligen Familie Podoviridae vorgeschlagen;[7][8][9][6][10][11] inklusive Gubaphagen.
Gattung „Chungbukvirus“[31] – war früher den Siphoviridae zugeordnet, weicht laut ICTV erheblich von diesen ab und hat Gemeinsamkeiten mit den Myoviridae; sie wird mit ihren Spezies vom ICTV aktuell nicht gelistet (wg. unvollständiger Genom-Daten).
Spezies „Thermocrinis Great Boiling Spring virus“ (TGBSV), Isolat GBS41 – Wirt: Thermocrinis (Aquificae), Fundort: Great Boiling Spring (GBS), Nevada[45][46]
David Veesler, Christian Cambillau: A Common Evolutionary Origin for Tailed-Bacteriophage Functional Modules and Bacterial Machineries. In: ASM Journals: Microbiology and Molecular Biology Reviews, Band 75, Nr. 3, 1. September 2011; S. 423–433; doi:10.1128/mmbr.00014, PMID 21885679, PMC 3165541 (freier Volltext) (englisch).
Maßgebliche Instanz ist das International Committee on Taxonomy of Viruses, viele der anderen Datenbanken haben derzeit (12. April 2022) dessen aktuellen Stand der Master Species List (MSL) #37, freigegeben Ende März 2022, noch nicht eingepflegt:
↑Andrey N. Shkoporov et al. (crAss-like phages Study Group): Proposal 2021.022B. Create one new order (Crassvirales) including four new families, ten new subfamilies, 42 new genera and 73 new species (Caudoviricetes). Stand: 13. Mai 2021.
↑Ajeng K. Pramono, Hirokazu Kuwahara, Takehiko Itoh, Atsushi Toyoda, Akinori Yamada, Yuichi Hongoh: Discovery and Complete Genome Sequence of a Bacteriophage from an Obligate Intracellular Symbiont of a Cellulolytic Protist in the Termite Gut. In: Microbes and Environments. 32. Jahrgang, Nr.2, 2017, ISSN1342-6311, S.112–117, doi:10.1264/jsme2.ME16175, PMID 28321010, PMC 5478533 (freier Volltext) – (englisch).
↑ abcY. Liu et al. (ICTV Archaeal Viruses Subcommittee): Proposal 2021.001A (zip:docx), PDF (via Universität Helsinki). Create three new orders and 14 new families in the class Caudoviricetes (Duplodnaviria, Uroviricota) for classification of archaeal tailed viruses. Oktober 2020.
↑B. Rihtman et al. (ICTV Bacterial Viruses Subcommittee): Proposal 2021.056B. Create one new family Naomiviridae including one new genus (Caudoviricetes)
↑
Sofia Medvedeva, Jiarui Sun, Natalya Yutin, Eugene V. Koonin, Takuro Nunoura, Christian Rinke, Mart Krupovic: [1] (zip:docx): Create one new order, ‘Atroposvirales’ and two new families, ‘Verdandiviridae’ and ‘Skuldviridae’ for classification of viruses of Asgardarchaeota. Oktober 2021.
↑ ab
Yue Su, Wenjing Zhang, Yantao Liang, Hongmin Wang, Yundan Liu, Kaiyang Zheng, Ziqi Liu, Hao Yu, Linyi Ren, Hongbing Shao, Yeong Yik Sung, Wen Jye Mok, Li Lian Wong, Yu-Zhong Zhang, Andrew McMinn, Min Wang: Identification and genomic analysis of temperate Halomonas bacteriophage vB_HmeY_H4907 from the surface sediment of the Mariana Trench at a depth of 8,900 m. In: ASM Journals: Microbiology Spectrum (Bacteriophages), 20. September 2023; doi:10.1128/spectrum.01912-23, PMID 37728551, PMC 10580944 (freier Volltext), ResearchGate (englisch). Siehe insbes. Fig. 1. Dazu:
↑
Jean-Gerard Tiraby, E. Tiraby, M. S. Fox: Pneumococcal bacteriophages. In: Virology Band 68, Dezember 1975, S. 566–569, doi:10.1016/0042-6822(75)90300-1, PMID 844.
↑Rubens López López: Streptococcus pneumoniae and its bacteriophages: one long argument. In: Int. Microbiol. 7. Jahrgang, Nr.3, 2004, S.163–71, PMID 15492930 (englisch).freier Volltext (PDF) Memento im Web-Archiv, 9. August 2017.
↑ ab
Marike Palmer, Brian P. Hedlund, Simon Roux, Philippos K. Tsourkas, Ryan K. Doss, Casey Stamereilers, Astha Mehta, Jeremy A. Dodsworth, Michael Lodes, Scott Monsma, Tijana Glavina Del Rio, Thomas W. Schoenfeld, Emiley A. Eloe-Fadrosh, David A. Mead: Diversity and Distribution of a Novel Genus of Hyperthermophilic Aquificae Viruses Encoding a Proof-Reading Family-A DNA Polymerase. In: Frontiers in Microbiology: Sec. Extreme Microbiology, Extremophiles: Microbial Genomics and Taxogenomics, Band 11, 12. November 2020, Nr. 583361 (eCollection 2020); doi:10.3389/fmicb.2020.583361, PMID 33281778, PMC 7689252 (freier Volltext), ResearchGate. Siehe insbesondere on image to zoom&p=PMC3&id=7689252_fmicb-11-583361-g001.jpg Fig. 1 und Supplement Data Sheet 1 (PDF).
↑
Rafael R. de la Haba, André Antunes,, Brian P. Hedlund: Editorial: Extremophiles: Microbial genomics and taxogenomics. In: Frontiers in Microbiology: Sec. Extreme Microbiology, Extremophiles: Microbial Genomics and Taxogenomics, Band 13, 2. August 2022; doi:10.3389/fmicb.2022.984632, PMID 35983330, PMC 9379316 (freier Volltext) (englisch).
↑
Luke J. McKay, Olivia D. Nigro, Mensur Dlakić, Karen M. Luttrell, Douglas B. Rusch, Matthew W. Fields, William P. Inskeep: Sulfur cycling and host-virus interactions in Aquificales-dominated biofilms from Yellowstone’s hottest ecosystems. In: Nature: The ISME Journal, Band 16, 14. Oktober 2021, S. 842–855; doi:10.1038/s41396-021-01132-4.





 Clostridium-Phage CPS2, Guelinviridae
Clostridium-Phage CPS2, Guelinviridae Lederbergvirus, Podoviren
Lederbergvirus, Podoviren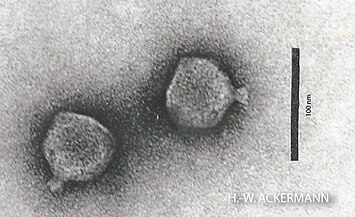 Bruynoghevirus, Podoviren
Bruynoghevirus, Podoviren Roseovarius-Virus RPP1, Schitoviridae
Roseovarius-Virus RPP1, Schitoviridae



{kind=link}