Arbacia punctulata

Arbacia punctulata est une espèce d'oursins du genre Arbacia, qui se rencontre dans l'Atlantique Ouest.
Description
[modifier | modifier le code]Arbacia punctulata est un oursin régulier de couleur sombre, généralement pourpre foncé mais pouvant aller du marron au noir en passant par différentes gammes de violet, de vert et de gris sombre. Les épines (« radioles ») sont lisses et peu effilées, épaisses et coniques ; elles sont densément réparties et généralement plus claires que le reste du corps. La coquille (« test ») arrondie est aplatie dorsalement, parfois presque hémisphérique, avec notamment une face orale presque parfaitement plate chez l'adulte mature, alors que le juvénile est globulaire. Le test de l'adulte dépasse rarement 5 cm de diamètre[1].
Son anus est entouré de quatre plaques madréporitiques, ce qui est une caractéristique du genre[1].
-

-
 Face aborale.
Face aborale. -
 Face orale.
Face orale. -
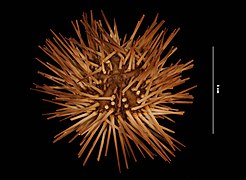 Spécimen de muséum
Spécimen de muséum -




_001.jpeg)

Répartition et habitat
[modifier | modifier le code]Son habitat naturel se situe dans l'océan Atlantique. On peut le trouver dans des eaux peu profondes depuis le Massachusetts jusqu'à Cuba et la péninsule du Yucatan, du Texas à la Floride dans le golfe du Mexique, la côte du Panama à la Guyane française et dans les Petites Antilles, le plus souvent dans les rochers, sur le sable, ou les plages coquillières[2].
Écologie et comportement
[modifier | modifier le code]Brouteur omnivore, charognard et prédateur opportuniste, A. punctulata consomme une grande variété de proies[3] bien que Karlson (1978)[4] le considère comme un carnivore pur.
Arbacia punctulata et l'Homme
[modifier | modifier le code]Interaction in situ
[modifier | modifier le code]Comme tous les oursins vivant à proximité de la surface, A. punctulata est souvent responsables de vives douleurs quand un baigneur marche dessus par inadvertance : ses épines ont tendance à se casser dans la plaie, ce qui les rend presque impossibles à enlever entièrement. Heureusement, il n'est pas venimeux, et ne présente pas de grand danger si la plaie est correctement désinfectée : le corps dissoudra les morceaux de silice en quelques semaines.
A. punctulata ne semble pas consommé de manière significative, et n'est pas récolté industriellement.
Utilisation scientifique
[modifier | modifier le code]Pendant plus d'un siècle, les embryologistes ont utilisé A. punctulata comme modèle expérimental. En effet, les oursins ont une fertilité extraordinaire, pouvant produire plusieurs millions d’œufs en une ponte, ce qui permet d'envisager des études quantitatives robustes. Par ailleurs, ces œufs peuvent être manipulés facilement dans les laboratoires de recherche, et sont transparents, ce qui permet de voir aisément les processus de développement qui s'y jouent. Enfin leurs œufs peuvent être facilement fécondés, se développer rapidement et de façon synchrone[5],[6].
Pendant des décennies, l'embryon d'oursin a été utilisé pour établir la théorie chromosomique de l'hérédité, la description des centrosomes, la parthénogenèse et la fécondation[7],[8],[9]. Les travaux de recherche au cours des trente dernières années, ont permis de comprendre des phénomènes tels que l'ARNm stable et le contrôle de la traduction génétique, l'isolement et la caractérisation du fuseau mitotique, et la réalisation que les principales protéines de structure de fuseau sont les microtubules[10],[11]. Les études sur les oursins ont fourni les premières preuves du rôle de l'actine dans les cellules non-musculaires[12],[13].
Elle est également un organisme modèle de la toxicité des sédiments marins[14],[15] et pour l'étude du sperme[16],[17].
Références taxinomiques
[modifier | modifier le code]- (en) WoRMS : espèce Arbacia punctulata (Lamarck, 1816)
- (fr) SeaLifeBase :
- (fr + en) ITIS : Arbacia punctulata (Lamarck, 1816)
- (en) Catalogue of Life : Arbacia punctulata (Lamarck, 1816) (consulté le )
- (en) Animal Diversity Web : Arbacia punctulata
- (en) NCBI : Arbacia punctulata (taxons inclus)
- (fr + en) EOL : Arbacia punctulata
Références
[modifier | modifier le code]- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Arbacia punctulata » (voir la liste des auteurs).
- Source : fiche sur le site de l'université de Jussieu.
- ↑ D. K. Serafy, 1979: Echinoids (Echinodermata: Echinoidea). Mem. Hourglass Cruises, 5: 1 – 120.
- ↑ D. T. Sharp & I. E. Gray, 1962: Studies on factors affecting the local distribution of two sea urchins, Arbacia punctulata and Lytechinus variegatus. Ecology, 43: 309 – 313.
- ↑ R. Karlson, 1978: Predation and space utilization patterns in a marine epifaunal community. J. Exp. Mar. Biol. Ecol., 31: 225 – 239.
- ↑ (en) O Rulon, « The modification of developmental patterns in Arbacia eggs with malonic acid », Anat. Rec., vol. 99, no 4, , p. 652 (PMID 18895450)
- ↑ (en) J Kanungo, « Prolonged incubation in seawater induces a DNA-dependent protein phosphorylation activity in Arbacia punctulata eggs », Biochem. Biophys. Res. Commun., vol. 294, no 3, , p. 667–71 (PMID 12056821, DOI 10.1016/S0006-291X(02)00539-9, lire en ligne)
- ↑ (en) PM Failla, « Recovery from division delay in irradiated gametes of Arbacia punctulata », Radiat. Res., vol. 25, no 2, , p. 331–40 (PMID 14295124, DOI 10.2307/3571975, lire en ligne)
- ↑ (en) MI Sachs, E Anderson, « A cytological study of artificial parthenogenesis in the sea urchin Arbacia punctulata », J. Cell Biol., vol. 47, no 1, , p. 140–58 (PMID 4327513, PMCID 2108410, DOI 10.1083/jcb.47.1.140, lire en ligne)
- ↑ (en) GL Kite, « The nature of the fertilization membrane of the egg of the sea urchin (Arbacia punctulata) », Science, vol. 36, no 930, , p. 562–564 (PMID 17812420, DOI 10.1126/science.36.930.562-a, lire en ligne)
- ↑ (en) AM Zimmerman, D Marsland, « Cell division: effects of pressure on the mitotic mechanisms of marine eggs (Arbacia punctulata) », Exp. Cell Res., vol. 35, , p. 293–302 (PMID 14195437, DOI 10.1016/0014-4827(64)90096-5)
- ↑ (en) A Scott, « A cytological analysis of the effects of cyanide and 4,6-dinitro-orthocresol on the mitotic phases in Arbacia punctulata », Biol. Bull., vol. 99, no 2, , p. 362–3 (PMID 14791535)
- ↑ (en) JH Henson, G Schatten, « Calcium regulation of the actin-mediated cytoskeletal transformation of sea urchin coelomocytes », Cell Motil., vol. 3, nos 5-6, , p. 525–34 (PMID 6420068, DOI 10.1002/cm.970030519)
- ↑ (en) J. Kabat-Zinn, RH Singer, « Sea urchin tube feet: unique structures that allow a cytological and molecular approach to the study of actin and its gene expression », J. Cell Biol., vol. 89, no 1, , p. 109–14 (PMID 6894447, PMCID 2111775, DOI 10.1083/jcb.89.1.109, lire en ligne)
- ↑ (en) Jäntschi L, Bolboaca SD, « A structural modelling study on marine sediments toxicity », Mar Drugs, vol. 6, no 2, , p. 372–88 (PMID 18728732, PMCID 2525494, DOI 10.3390/md20080017)
- ↑ (en) A Rudolph, P Medina, C Urrutia, R Ahumada, « Ecotoxicological sediment evaluations in marine aquaculture areas of Chile », Environ Monit Assess, vol. 155, nos 1-4, , p. 419–29 (PMID 18633720, DOI 10.1007/s10661-008-0444-x)
- ↑ (en) FR Lillie, « The Fertilizing Power of Sperm Dilutions of Arbacia », Proceedings of the National Academy of Sciences U.S.A., vol. 1, no 3, , p. 156–60 (PMID 16575966, PMCID 1090763, DOI 10.1073/pnas.1.3.156)
- ↑ (en) MV Inamdar, T Kim, YK Chung, et al., « Assessment of sperm chemokinesis with exposure to jelly coats of sea urchin eggs and resact: a microfluidic experiment and numerical study. », J. Exp. Biol., vol. 210, no Pt 21, , p. 3805–20 (PMID 17951422, DOI 10.1242/jeb.005439, lire en ligne)