フタマタケカビ
| フタマタケカビ | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| ||||||||||||||||||
| 分類(目以上はHibbett et al. 2007) | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Syzygites Ehrenb. 1832 | ||||||||||||||||||
| 種 | ||||||||||||||||||
|
本文参照 |
フタマタケカビ Syzygites はケカビ目に属するカビ。様々なキノコに発生し、胞子嚢柄が繰り返し2叉分枝する。
特徴
[編集]本属の特徴は直立した胞子嚢柄が複数回にわたって2叉分枝を行い、それぞれの枝の先端に多数の胞子を含む柱軸を持つ胞子嚢を形成すること、自家和合生でケカビ型の接合胞子嚢を作ること、である[1]。
以下、やや詳しく示す。本属には単一の種、S. megarocarpus Ehrenb. 1832 のみが知られており、以下はこの種に基づいて記す[2]。
栄養体
[編集]通常の培地で培養が可能であり、ジャガイモショ糖寒天培地上で23~25℃で培養した際の成長は2~3日でペトリ皿の縁に達するほどに早い。コロニーは綿毛状になり、多くの気中菌糸を出し、淡黄褐色を呈する。なお、成長は32℃まで認められる。
無性生殖
[編集]無性生殖は多胞子の胞子嚢による。培養下における胞子嚢胞子の形成は3日目以降に生じる。胞子嚢柄は長さ5cmまでになる。その径は30~55μmで、隔壁を有し、無色透明、表面は滑らか。ただし少なくともその基部に近い部分では縦筋があったり凹凸があったりしている[1]。胞子嚢柄は2叉に繰り返して分枝している。その分枝は最大で6回[1]。後に青みを帯びた色に変わる。胞子嚢は始め黄色で後に明るい灰色から暗い灰色に変わり、肉眼でも認められる。胞子嚢は球形からやや不規則な球形をしており、その径は50~150μm[3]。柱軸は亜球形から不規則な球形で径30~200μm。胞子嚢胞子は球形から不規則な球形、あるいは卵形で長さ5~35μm、表面には粒状の突起が並び透明から淡褐色となっている。
-
 胞子嚢柄の様子
胞子嚢柄の様子 -
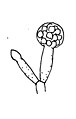 胞子嚢と柱軸
胞子嚢と柱軸


有性生殖
[編集]有性生殖は接合胞子の形成による。自家和合性であり、単一の株でもよく接合胞子を形成する。分化した接合胞子嚢柄を持ち、接合胞子柄は栄養菌糸から出て立ち上がり、二叉分枝し、その枝の間で接合胞子を生じる[4]。接合子嚢は向かい合う形で接合し、いわゆるケカビ型である。接合胞子嚢支持柄は同型で丸く膨らむ[4]。接合胞子嚢は当初は透明で、後に成熟すると赤褐色か暗褐色に変化し、その形は球形から亜球形で、大きさは160~230μm×190~270μm程、表面は粒状突起に被われ、その外壁は肥厚して厚さ5μmほどになる。時として接合によらずに形成される単為接合胞子嚢が作られる[4]。
-
 配偶子嚢が接合する段階
配偶子嚢が接合する段階 -
 完成した接合子嚢
完成した接合子嚢 -
 単為接合子嚢
単為接合子嚢



生態
[編集]本種は生きたキノコの子実体上に出現するものであり、従って菌寄生菌とされる。ただし上記のように純粋培養が可能であり、しかもごく標準的な培地でよく生育する。従って条件的寄生菌と呼ばれるものに相当する。いわゆる菌寄生菌はその栄養の型によって biotrophic とnecrotrophic という2つの型が認められ、前者は宿主の生きた細胞から栄養を求め、その多くは宿主の範囲が限定される[5]。後者は殺生ともいい、宿主細胞を殺した後に栄養を吸収し、この型のものは宿主の対象が幅広いことが多い。本種は後者に当たると考えられる。ちなみに宇田川他(1978)では菌寄生とも書いておらず『腐った肉質のキノコ上』に見られる、とある[4]。
本種の寄生対象の範囲に関しては、例えば北アメリカ、イリノイ州での報告では11科に渡る23種が挙げられている[1]。内容としてはハラタケ目が大部分であるが腹菌類が2種(ノウタケ属とホコリタケ属)含まれ、更に多孔菌が1種(霊芝)、更に子嚢菌のアミガサタケ属の1種までが含まれ、宿主の選択性が低いことがわかる。韓国からは野生のキノコでの報告として、マツタケを含む5種類が本菌に寄生を受けていることが報告されている[6]。
分布
[編集]北半球に広く分布するもので、ヨーロッパと南北アメリカ[7]、それに東アジア[8]から知られる。日本でも広く見られる[4]。
類似のもの
[編集]キノコに寄生するケカビ目の菌としては本属の他にタケハリカビ Spinellus、ディクラノフォラ Dranophora などが挙げられる。前者は胞子嚢柄が全く分枝せずに伸びて、まるでキノコの傘が針山のようになるもので、後者は大型の胞子嚢を付ける枝と、それとは別に細かく分枝した枝に小胞子嚢をつけるもので、キノコの表面が綿毛に被われたような姿になる。なお、この2者共に低温を好む菌で、20℃以下で出現するものである。
形態の上からは、胞子嚢柄が多数分枝を出す例にはシャジクケカビ Actinomucor があるが、この場合、頂端に大きな胞子嚢を付け、その側枝が枝分かれするものであり、また繰り返し2叉分枝することもない。胞子嚢柄が2叉分枝を繰り返すのはディコトモクラディウム Dichotomocladium などの例もあるが、枝の先端に胞子嚢を付けるわけではない。エダケカビ Thamnidium やサムノスチルム Thamnostyrum などにも小胞子嚢が2叉分枝した枝につく例があるが、これらは大型の胞子嚢とは異なり、その壁が壊れて胞子散布をすることは少ない。かつてケカビ目に含めたエダカビ Piptocephalis は先端に向けて二叉分枝を繰り返す枝を作るが、この場合、枝先につくのが多数の分節胞子嚢である。
スポロディニエラ Sporodiniella は胞子嚢柄の先端近くで散形に分枝が出て、その先端に大きい胞子嚢を付ける点で似ているが、胞子嚢の直下に不実の棘を持つ。またこの菌は昆虫から出現する。ちなみにこの菌の学名は本属の異名である Sporodinia に基づく。
分類
[編集]古典的な分類体系では本属は大型で柱軸がある胞子嚢のみを作る点からケカビ科 Mucoraceae とし[9]、あるいは条件的寄生菌であることからディクラノフォラなどとまとめてディクラノホラ科 Dicranophraceae とする[10]ことなどが行われてきたが、そのような主として形態に基づく体系は分子系統の情報などから大きく見直されている。Hoffmann et al.(2013)では本属はクモノスカビ属 Rhizopus と同一のクレードに含まれ、これをクモノスカビ科 Rhizopodaceae とする、との判断となっている。このクレードの中には更に3つのクレードがあるが、本属が含まれるクレードにはもっとも普通に見られるクモノスカビである R. stronifer と、それに上述のスポロディニエラが含まれる。これらは共通の性質として、比較的高温耐性があること、他生物への寄生的傾向があることなどが挙げられている[11]。
利害
[編集]本種は菌寄生菌であり、キノコを宿主とする。この宿主には食用などに利用される種が幾つも含まれている。上述のイリノイ州での調査ではハラタケ属 Agaricus のツクリタケ A. bisporus 、ハラタケ A. campestris、カワリハラタケ(いわゆるアガリクス) A. subrufescens、それにシイタケ Lentinus edodes、レイシ Ganoderma lucidum が含まれている。また韓国ではマツタケ Tricholoma matsutake に本種が寄生することも報告されている[6]。ただしこれらいずれに関しても本種の出現率が高くないためか、重要な脅威とは見なされていない模様である[8]。
菌類学史の上で
[編集]菌類の性に関する研究史の上では、本属の唯一の種である M. megalocarpus は重要な役割を果たしてきた[12]。本菌は菌類に於いて性に関して初めて報告された(1820年)もので、また自家不和合性 Heterothallic に対する形で自家和合性 Homothallic という概念が適用された最初の菌(1904年)であり、接合胞子 Zygospore という語が当てられたのも本種が最初であった。
この時期、キノコやカビの胞子が生殖細胞であることは理解されていたが、そこに有性的な働きがあるかどうかは明らかにされていなかった[13]。植物の花との関連で雄しべや雌しべに当たるものがキノコのどこかにないかという議論がなされていた時期であり、シスチジアに性的役割があるのではないかといった議論がなされていたりもしている。そんな中で本菌は1818年にエーレンベルクによって発見され、彼はその2年後にこの菌が接合胞子を形成することを見いだし、それをはっきりした図を含めて発表した。これに先立ってアオミドロにおいて接合が観察され(1782年)、これが有性的な活動であることも認められ(1803年)ていたため、彼はこれと比較することで検討し、本菌がケカビに類する菌類であり、その有性的な活動であるとした。その重要性は当時認められず、1860年代になってド・バリらが本菌でこれを再確認して、これを接合であり、有性生殖の活動であると認め、その胞子にアオミドロで認められた名である接合胞子という語を当てた。この直後、クモノスカビ属やケカビ属の接合胞子も確認された。なおこの前後に卵菌類の生殖器官に関しても発見が相次ぎ、またド・バリは子嚢菌の有性生殖の構造の発見にも与っている。いわゆるキノコの有性生殖の仕組みについて研究が進むようになったのはそれ以降のことである。
出典
[編集]- ^ a b c d Kovacs & Sundberg(1999)
- ^ 以下、主としてKa et al.(1999)
- ^ 以下、胞子嚢に関してはKovacs & Sundberg(1999)
- ^ a b c d e 宇田川他(1978),p.307
- ^ 以下、Ka et al.(1999)
- ^ a b Ka et al.(2000)
- ^ Zycha et al.(1969),p.73
- ^ a b Ka et al.(1999)
- ^ Zycha et al.(1969)
- ^ Alexopoulos et al.(1996)
- ^ Hoffmann et al.(2013),p.72
- ^ 以下、Idnurum(2011)
- ^ 以下、エインズワース/小川(2010),p.121
参考文献
[編集]- 宇田川俊一他、『菌類図鑑(上)』、(1978)、講談社
- G. C. エインズワース/小川眞訳、『キノコ・カビの研究史――人が菌類を知るまで』、(2010)、京都大学学術出版会
- C.J.Alexopoulos,C.W.Mims, & M.Blackwell,INTRODUCTORY MYCOLOGY 4th edition,1996, John Wiley & Sons,Inc.
- Zycha H.,R. Shiepmann, G. Linnemann, 1969. Mucorales eine eschreibung aller Gattungen und Alten dieser Pilzgruppe. J. Carrmer, Lehre, Germany. 355pp.
- Kang-Hyeon Ka et al. 1999. Syzygites megalocarpus (Mucorales): A necrotrophic mycoparasite of Tricholoma matutake. The Korean Journal of Mycology Vol.17 No.5 p.345-348
- Kang-Hyeon Ka et al. 2000. Occurrence of Syzygites megalocarus on Wild Mushroom and its Cultural Characterisutics. Jour. Korean For. Sci. 89(5): p.691-696.
- R. K. Kovacs & W. J. Sundverg, 1999. Syzygites megalocarpus (Mucorales, Zygomycetes) in Illinois. Transactions of the Illinois State Academy of Science Vol.92 3&4: p.181-190.
- Alexander Idnurm, 2011. Sex Determination in the First-Described Sexual Fungus. Eucariotic Cell Nov. 2011: p.1485-1491.