Dinosaur Park-formatie
De Dinosaur Park-formatie is de bovenste afzetting van de Belly River Group (ook bekend als de Judith River Group), een belangrijke geologische eenheid in het zuiden van Alberta. Het werd afgezet tijdens het Campanien van het Laat-Krijt, tussen ongeveer 76,5 en 74,4 miljoen jaar geleden. Het werd afgezet in alluviale en kustvlakte-omgevingen en wordt begrensd door de niet-mariene Oldman-formatie eronder en de mariene Bearpaw-formatie erboven.
De Dinosaur Park-formatie bevat dichte concentraties dinosauriërskeletten, zowel in anatomisch verband liggend als onsamenhangend, die vaak worden gevonden met bewaard gebleven overblijfselen van zachte weefsels. Overblijfselen van andere dieren zoals vissen, schildpadden en krokodilachtigen, evenals plantenresten, zijn ook overvloedig aanwezig. De formatie is vernoemd naar het Dinosaur Provincial Park, een UNESCO-werelderfgoedlocatie waar de formatie goed zichtbaar is in de badlands die de Red Deer River flankeren.
Geologische locatie
[bewerken | brontekst bewerken]De Dinosaur Park-formatie is samengesteld uit sedimenten die zijn afgeleid van de erosie van de bergen in het westen. Het werd afgezet op een alluviale tot kustvlakte door riviersystemen die oostwaarts en zuidoostwaarts stroomden naar de Bearpaw Sea, een grote binnenzee die deel uitmaakte van de Western Interior Seaway. Die zee overspoelde geleidelijk de aangrenzende kustvlakte, waardoor de mariene leisteen van de Bearpaw-formatie bovenop de Dinosaur Park-formatie werd afgezet.
De Dinosaur Park-formatie is ongeveer zeventig meter dik in Dinosaur Park. Het onderste deel van de formatie werd neergelegd in riviergeulomgevingen en bestaat voornamelijk uit fijn- tot mediumkorrelige, kruiselings ingebedde zandstenen. Het bovenste gedeelte, dat werd afgezet in oever- en overstromingsvlakten, bestaat voornamelijk uit massieve tot gelamineerde, organisch-rijke moddersteen met overvloedige wortelsporen en dunne bedden van bentoniet. De Lethbridge Coal Zone, die bestaat uit verschillende lagen laagwaardige steenkool die zijn ingebed in moddersteen en siltstones, markeert de top van de formatie.
De sedimenten van de Dinosaur Park-formatie zijn vergelijkbaar met die van de onderliggende Oldman-formatie en ze waren oorspronkelijk opgenomen in die formatie. De twee formaties zijn echter gescheiden door een regionale disconformiteit en onderscheiden zich door petrografische en sedimentologische verschillen. Bovendien zijn gearticuleerde skeletresten en botbedden zeldzaam in de Oldman-formatie, maar overvloedig in de Dinosaur Park-formatie.
Biostratigrafie
[bewerken | brontekst bewerken]De Dinosaur Park-formatie kan worden onderverdeeld in ten minste twee verschillende fauna's. Het onderste deel van de formatie wordt gekenmerkt door de overvloed aan Corythosaurus en Centrosaurus. Deze groep soorten wordt hoger in de formatie vervangen door een andere ornithischische fauna gekenmerkt door de aanwezigheid van Lambeosaurus en Styracosaurus. Het verschijnen van verschillende nieuwe, zeldzame soorten Ornithischia helemaal bovenaan de formatie kan erop wijzen dat een derde aparte fauna de tweede had vervangen tijdens de overgang naar jongere, niet-Dinosaur Park-sedimenten, terwijl tegelijkertijd een binnenzee het land binnendringt, maar er zijn hier minder overblijfselen. Een niet nader genoemde pachyrhinosaurïer, Vagaceratops irvinensis en Lambeosaurus magnicristatus komen mogelijk vaker voor in deze derde fauna.
De onderstaande tijdlijn volgt een synthese gepresenteerd door Fowler (2017) met aanvullende informatie van Arbor et alii 2009, Evans et al. 2009 en Penkalski, 2013. Megaherbivore Assemblage Zones (MAZ) volgen gegevens gepresenteerd door Mallon et alii, 2012.

Amfibieën
[bewerken | brontekst bewerken]Overblijfselen van de volgende amfibieën zijn gevonden in de formatie:
Caudata (salamanders)
- Habrosaurus prodilatus
- Lisserpeton
- Opisthotriton kayi
- Scapherpeton tectum
- naamloze caudata
- Twee onbepaalde caudata
Salienta (kikkers)
- Twee naamloze salientiden
- Tyrrellbatrachus brinkmani
- Hensonbatrachus kermiti
Albanerpetontidae (uitgestorven, salamanderachtige amfibieën)
- Albanerpeton gracilis
Dinosauriërs
[bewerken | brontekst bewerken]Overblijfselen van de volgende dinosauriërs zijn gevonden in de formatie:
Ornithischia
[bewerken | brontekst bewerken]Overblijfselen van de volgende ornithischiërs zijn gevonden in de formatie:
Ankylosauriërs
[bewerken | brontekst bewerken]| Ankylosauriërs van de Dinosaur Park Formation | |||||
|---|---|---|---|---|---|
| Geslacht | Soort | Stratigrafische positie | Materiaal | Opmerking | Afbeelding |
| Anodontosaurus | A. inceptus | Midden, 75.6 miljoen jaar geleden | Twee schedels met tanden, onderkaak, gedeeltelijk geprepareerd skelet, beide cervicale halfringen en osteodermen. | Een ankylosaurine ankylosauride |  |
| Dyoplosaurus | D. acutosquameus | Vroeg, 76.5 miljoen jaar geleden | Een gedeeltelijke schedel en skelet inclusief bekken, staart en achterpoot met voet en osteodermen. | Een ankylosaurine ankylosauride | 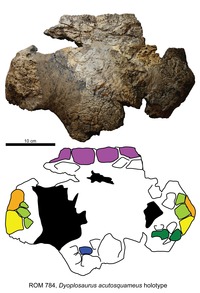 |
| Edmontonia | E. rugosidens | Vroeg, 76.5-75.9 miljoen jaar geleden | Een gedeeltelijk skelet inclusief een schedel, ruggenwervels, proximale, distale caudale, ribben, opperarmbeen, ellepijp, spaakbeen, manus, fragmenten van het bekken, scheenbeen, kuitbeen?, osteodermen; voorste helft van een in verband liggend skelet met in situ osteodermen en gepaarde eerste mediale schubben. | Een nodosaurine nodosaurid also known from the Horseshoe Canyon Formation and Two Medicine Formation |  |
| Euoplocephalus | E. tutus | Vroeg tot Midden, ~76.4-75.6 miljoen jaar geleden | Vier schedels, onderkaak, halswervels, ruggenwervels, ribben, schouderbladen, opperarmbeen, spaakbeen, ellepijp, middenhandsbeentjes, kootjes, darmbeen, zitbeen, dijbeen, scheenbeen, gedeeltelijke voet, heiligbeen, cervicale halfringen en osteodermen. | Een ankylosaurine ankylosauride | 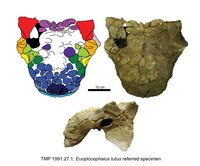 |
| Panoplosaurus | P. mirus | Midden, 75.6 miljoen jaar geleden | Schedel met onderkaak, losse tanden, halswervels, ruggenwervels, sacrale wervels, halsribben, rugribben, scapulocoracoïde, opperarmbeen, manus, scheenbeen, kuitbeen, verbeende intersternale plaat, een paar verbeende processus xiphoïdei, voet en in situ osteodermen. | Een nodosaurine nodosauride |  |
| Platypelta | P. coombsi | Vroeg, 77.5-76.5 miljoen jaar geleden | Een goed geconserveerde schedel, kaken, tanden, hals- en ruggenwervels, ribben, compleet bekken, beide scapulocoracoïden, zowel opperarmbeenderen als spaakbeenderen, beide cervicale halfringen en osteodermen. | Een ankylosaurine ankylosauride |  |
| Scolosaurus | S. cutleri | Vroeg, 76.5 miljoen jaar geleden of meer | Een bijna compleet skelet, een schedel, hals-, staart- en ruggenwervels, ribben, schouderblad, ravenbeksbeen, opperarmbeen, spaakbeenderen, darmbeen, zitbeen, dijbeen, scheenbeen, kuitbeen, een cervicale halfring en osteodermen. | Een ankylosaurine ankylosauride waarvan kort werd gedacht dat hij synoniem was aan Euoplocephalus. Het kwam mogelijk uit de bovenste lagen van de onderliggende Oldman Formation. |  |
| S. thronus | Laat, 75 miljoen jaar geleden | Een gedeeltelijk skelet inclusief een schedel, ruggenwervels, volledig synsacrum, sacrale ribben, staartwervels, schouderblad, gedeeltelijke darmbeenderen, opperarmbeen, cervicale halfringen, osteodermen en huidafdrukken. | Een ankylosaurine ankylosauride | ||
Ceratopiden
[bewerken | brontekst bewerken]Een naamloos Pachyrhinosaurus-achtig taxon is uit de formatie geborgen.
| Ceratopiërs van de Dinosaur Park Formation | |||||
|---|---|---|---|---|---|
| Geslacht | Soort | Stratigraphische positie | Materiaal | Opmerking | Afbeeldingen |
| Centrosaurus | C. apertus | Midden, 76.2-75.5 miljoen jaar geleden | Vijftien schedels, verschillende skeletten, allemaal volwassen; overvloedig beenderbedmateriaal met zeldzame juvenielen en subadulten. C. nasicornis kan een synoniem zijn. | Een centrosaurine ceratopide |  |
| Chasmosaurus | C. belli | Midden, 76–75.5 miljoen jaar geleden | Twaalf schedels, verschillende skeletten. | Een chasmosaurine ceratopide |  |
| C. russelli | Vroeg, 76.5-76 miljoen jaar geleden | Zes volledige of gedeeltelijke schedels. | |||
| Mercuriceratops | M. gemini | Vroeg, ~77 miljoen jaar geleden | Een apomorf squamosum. | Een chasmosaurine ceratopide |  |
| Monoclonius | M. lowei | Een dubieuze centrosaurine ceratopide. Mogelijk synoniem met Centrosaurus | |||
| Pentaceratops | P. aquilonius | Bovenste, 74.8 miljoen jaar geleden | twee schildfragmenten. | Een dubieuze chasmosaurine ceratopide die dezelfde soort kan zijn als Spiclypeus shipporum |  |
| Spinops | S. sternbergorum | Vroeg, 76.5 miljoen jaar geleden | Gedeeltelijk wandbeen, gedeeltelijk dentarium, niet-identificeerbare ledemaatfragmenten, gedeeltelijke schedel en gedeeltelijk rechtersquamosum. | Een centrosaurine ceratopide. Hpj kan zelfs afkomstig zijn uit de bovenste Oldman Formation. |  |
| Styracosaurus | S. albertensis | Laat, 75.5-75.2 miljoen jaar geleden | Twee schedels, drie skeletten, aanvullend materiaal in beenderbedden. | Een centrosaurine ceratopide |  |
| Unescoceratops | U. koppelhusae | Gedeeltelijke onderkaak. | Een leptoceratopide tussen de één en twee meter lang en minder dan 91 kilogram zwaar. Zijn tanden waren de rondste van alle leptoceratopiden. | ||
| Vagaceratops | V. irvinensis | Laat, 75 miljoen jaar geleden | Drie schedels, skelet zonder staart. | Een chasmosaurine ceratopide soort die eerder werd geclassificeerd als een soort van Chasmosaurus. |  |
Euornithopoden
[bewerken | brontekst bewerken]Ten minste één onbepaald exemplaar van de Thescelosauridae is uit de formatie geborgen.
In een recensie uit 2001 van eierschalen van hadrosauriërs en materiaal van jongen uit de Dinosaur Park Formation, concludeerden Darren H. Tanke en M.K. Brett-Surman dat hadrosauriërs nestelden in zowel het oude hoogland als de laaglanden van de afzettingsomgeving van de formatie. De hooggelegen broedgebieden hadden misschien de voorkeur van de minder gebruikelijke hadrosauriërs, zoals Brachylophosaurus of Parasaurolophus. De auteurs waren echter niet in staat om vast te stellen welke specifieke factoren de keuze van de nestplaats vormden in de hadrosauriërs van de formatie. Ze suggereerden dat gedrag, voeding, bodemgesteldheid en competitie tussen dinosauriërsoorten mogelijk allemaal van invloed waren op de plek waar hadrosauriërs nestelden.
Fragmenten van minder dan een centimeter van hadrosauriër-eierschaal met kiezelsteenstructuur zijn gerapporteerd uit de Dinosaur Park Formation. Deze eierschaal is vergelijkbaar met de eierschaal van de hadrosauriër van Devil's Coulee in het zuiden van Alberta, evenals die van de Two Medicine en Judith River Formations in Montana, Verenigde Staten. Hoewel aanwezig, is de eierschaal van dinosauriërs zeer zeldzaam in de Dinosaur Park Formation en wordt deze alleen gevonden op twee verschillende microfossiele locaties. Deze vindplaatsen onderscheiden zich door grote aantallen schelpdieren en andere minder vaak voorkomende ongewervelde dieren, zoals Unionidae en slakken. Deze associatie is geen toeval, aangezien de ongewervelde schelpen langzaam zouden zijn opgelost en voldoende basisch calciumcarbonaat zouden hebben afgegeven om de eierschalen te beschermen tegen van nature voorkomende zuren die ze anders zouden hebben opgelost en fossilisatie zouden hebben voorkomen.
In tegenstelling tot eierschaalfossielen komen de overblijfselen van zeer jonge hadrosauriërs eigenlijk vrij algemeen voor. Darren Tanke heeft opgemerkt dat een ervaren verzamelaar op één dag meerdere juveniele hadrosauriërs kan ontdekken. De meest voorkomende overblijfselen van jonge hadrosauriërs in de Dinosaur Park Formation zijn dentaria, botten van ledematen en voeten, evenals wervelcentra. Het materiaal vertoonde weinig of geen van de slijtage die het gevolg zou zijn van transport, wat betekent dat de fossielen werden begraven in de buurt van hun punt van herkomst. Beenderbedden 23, 28, 47 en 50 zijn productieve bronnen van jonge hadrosauriërresten in de formatie, vooral beenderbed 50. Van de botten van juveniele hadrosauriërs en fossiele eierschaalfragmenten is niet bekend dat ze samen met elkaar bewaard zijn gebleven, ondanks dat beide aanwezig zijn in de formatie.
| Euornithopoda van de Dinosaur Park Formation | |||||
|---|---|---|---|---|---|
| Geslacht | Soort | Stratigraphische positie | Materiaal | Opmerking | Afbeeldingen |
| Corythosaurus | C. casuarius | Vroeg midden, 76.5-75.5 miljoen jaar geleden | Ongeveer tien in verband liggende schedels en bijbehorende postcrania, tien tot vijftien in verband liggende losse schedels, losse schedelelementen, juveniel tot volwassen. | Een lide van de Lambeosaurini, een lambeosaurine hadrosauriër |  |
| Gryposaurus | G. notabilis | Vroeg, 76.2-76 miljoen jaar geleden | Ongeveer tien complete schedels, twaalf fragmentarische schedels, bijbehorende postcrania. | Een lid van de Kritosaurini, een saurolophine hadrosauriër |  |
| Lambeosaurus | L. lambei | Laat, 75.5-75 miljoen jaar geleden | Ongeveer zeven in verband liggende schedels met bijbehorende postcrania, mogelijk tien in verband liggende losse schedels, losse schedelelementen, juveniel tot volwassen. | ||
| L. magnicristatus | Laat/Bearpaw Formation, 74.8 miljoen jaar geleden | Twee complete schedels, één met bijbehorende in verband liggende postcrania. |  | ||
| Parasaurolophus | P. walkeri | Vroeg, 76.5-75.3 miljoen jaar geleden | Complete schedel en postcraniaal skelet. | Een lid van de Parasaurolophini, een lambeosaurine hadrosauriër. |  |
| Prosaurolophus | P. maximus | Laat, 75.5 – 74.8 miljoen jaar geleden | Twintig tot vijfentwintig individuen, waaronder ten minste zeven in verband liggende schedels en bijbehorende postcrania. | Een lid van de Saurolophini, een saurolophine hadrosauriër |  |
Pachycephalosauriërs
[bewerken | brontekst bewerken]| Pachycephalosauriërs van de Dinosaur Park Formation | |||||
|---|---|---|---|---|---|
| Geslacht | Soort | Stratigraphische positie | Materiaal | Opmerking | Afbeeldingen |
| Foraminacephale | F. brevis | Ook aanwezig in de Oldman Formation | Frontoparietale koepel, verschillende andere schedelfragmenten, waaronder juveniel en subadult materiaal. | Ooit gezien als een soort van Stegoceras | 
|
| Gravitholus | G. albertae | Frontopariëtale koepel. |  | ||
| Hanssuesia | H. sternbergi | Vroeg, ook aanwezig in de Oldman Formation en Judith River Formation |  | ||
| Stegoceras | S. validum | Specimina inclusief frontopariëtale koepel. |  | ||
| Microcephale | Een nomen nudum. | ||||
Theropoden
[bewerken | brontekst bewerken]In de Dinosaur Park Formation zijn kleine theropoden zeldzaam vanwege de neiging van hun dunwandige botten om te breken of slecht bewaard te blijven. Kleine botten van kleine theropoden die door grotere werden belaagd, zijn mogelijk in hun geheel doorgeslikt en verteerd. In deze context was de ontdekking van een kleine theropode dinosauriër met bewaarde tandafdrukken bijzonder waardevol. Mogelijke onbepaalde avimimide resten zijn bekend uit de formatie.
Ornithomimiden
[bewerken | brontekst bewerken]| Ornithomimiden van de Dinosaur Park Formation | |||||
|---|---|---|---|---|---|
| Geslacht | Soort | Stratigraphische positie | Materiaal | Opmerking | Afbeeldingen |
| Ornithomimus | O. sp. | Een ornithomimide, mogelijk een soort van Struthiomimus. |  | ||
| Qiupalong | Q. sp. | Verschillende specimina | Een ornithomimide, mogelijk een radiatie van dit geslacht uit Azië. |  | |
| Rativates | R. evadens | Holotype ROM 1790 | Een ornithomimide, vroeger gezien als een exemplaar van Struthiomimus. |  | |
Oviraptorosauriërs
[bewerken | brontekst bewerken]| Oviraptorosauriërs van de Dinosaur Park Formation | |||||
|---|---|---|---|---|---|
| Geslacht | Soort | Stratigraphische positie | Materiaal | Opmerking | Afbeeldingen |
| Caenagnathus | C. collinsi | Onderkaak, type-exemplaar | Een caenagnathide die in grootte wedijverde met Anzu. |  | |
| Chirostenotes | C. pergracilis | Verschillende fragmentarische exemplaren, type-exemplaar | Een middelgrote caenagnathide. |  | |
| Citipes | C. elegans | Verschillende fragmentarische exemplaren, type-exemplaar | Kleinste caenagnathide uit de formatie. |  | |
| Macrophalangia | M. canadensis | Jonger synoniem van Chirostenotes pergracilis | |||
Paraven
[bewerken | brontekst bewerken]Een nieuw taxon van troödontiden dat uitsluitend op tanden is gebaseerd, is bekend uit het bovenste deel van de formatie.
| Paraves van de Dinosaur Park Formation | |||||
|---|---|---|---|---|---|
| Geslacht | Soort | Stratigraphische positie | Materiaal | Opmerking | Afbeeldingen |
| cf. Baptornis | Onbepaald | Een hesperornithiforme vogel | |||
| cf. Cimolopteryx | Onbepaald | Gedeeltelijk ravenbeksbeen | Een mogelijke charadriiforme vogel | ||
| Dromaeosaurus | D. albertensis | Diverse exemplaren en tanden, type-exemplaar | Een dromaeosauride |  | |
| Hesperonychus | H. elizabethae | Bekkenbeenderen en gedeeltelijke tenen en klauwen, type-exemplaar | Een microraptorine dromaeosauride, ook gevonden in de Oldman Formation |  | |
| Latenivenatrix | L. mcmasterae | Bekkenbeenderen, schedelfragmenten, type-exemplaar | Een grote troödontide | .png) | |
| cf. Palintropus | Unnamed | Gedeeltelijke schoudergordels | Een ambiortiforme vogel | ||
| cf. Paronychodon | cf. P. lacustris | Tanden | Een onbepaalde maniraptoor, ook gevonden in de Judith-rivier | ||
| cf. Pectinodon | Onbepaalde troödontide | Tanden | Een troödontide | ||
| Polyodontosaurus | P. grandis | Dentarium, type-exemplaar | Nomen dubium. Mogelijk synoniem met Latenivenatrix. | ||
| Richardoestesia | R. gilmorei | Onderkaak, type-exemplaar | Een dromaeosauride | ||
| R. isosceles | Tanden | ||||
| Saurornitholestes | S. langstoni | Incompleet skelet en tanden, type-exemplaar. Een aan Saurornitholestes toegewezen dentarium werd ontdekt dat de tandafdrukken van een jonge tyrannosauriër bewaarde. | Een dromaeosauride |  | |
| Stenonychosaurus | S. inequalis | Bijna compleet skelet en andere gedeeltelijke skeletten, type-exemplaar | Een troödontide, ooit gezien als identiek aan Troodon formosus |  | |
Tyrannosauriden
[bewerken | brontekst bewerken]| Tyrannosauriden van de Dinosaur Park Formation | |||||
|---|---|---|---|---|---|
| Geslacht | Soort | Stratigrafische positie | Materiaal | Opmerking | Afbeeldingen |
| Daspletosaurus | Naamloze soort | Midden-Laat, 75.6-75 miljoen jaar geleden | Meerdere exemplaren | Een tyrannosaurine tyrannosauride, ook aanwezig in de Bearpaw Formation. | 
|
| Gorgosaurus | G. libratus | Vroeg-Midden, 76.6-75.1 miljoen jaar geleden | Talrijke exemplaren, type-exemplaar | Een albertosaurine tyrannosauride wiens fossielen zijn opgegraven in de Judith River Formation en mogelijk de Two Medicine Formation. Het was de meest voorkomende grote carnivoor in het gebied. |  |
Andere reptielen
[bewerken | brontekst bewerken]Choristoderes
[bewerken | brontekst bewerken]Choristoderes, of champsosauriërs, waren in het water levende reptielen. Kleine exemplaren leken op hagedissen, terwijl grotere soorten oppervlakkig gezien op krokodilachtigen leken. Overblijfselen van de volgende Choristoderes zijn gevonden in de formatie:
- Champsosaurus (ten minste drie soorten)
- Cteniogenys sp. cf. antiquus (mogelijk een ander geslacht)
Crocodylia
[bewerken | brontekst bewerken]Overblijfselen van de volgende Crocodylia zijn gevonden in de formatie:
- Albertochampsa
- Leidyosuchus
- Ten minste een naamloos taxon
Hagedissen
[bewerken | brontekst bewerken]Overblijfselen van de volgende hagedissen zijn gevonden in de formatie:
- Helodermatiden
- Labrodioctes
- Necrosauriden
- Parasaniwa
- Teiiden
- Glyptogenys
- Socognathus
- Varaniden
- Palaeosaniwa
- Xenosauriden
- ?Exostinus
Plesiosauriërs
[bewerken | brontekst bewerken]Overblijfselen van de volgende plesiosauriërs zijn gevonden in de formatie:
- Fluvionectes
- onbepaalde polycotyliden (kortere hals)
Pterosauriërs
[bewerken | brontekst bewerken]Overblijfselen van de volgende pterosauriërs zijn gevonden in de formatie:
- Cryodrakon (bekend van kleine en grote exemplaren)
- Een naamloze niet-Azhdarchide pterosauriër
Schildpadden
[bewerken | brontekst bewerken]Overblijfselen van de volgende schildpadden zijn gevonden in de formatie:
- Adocus
- Apalone
- Aspideretoides (drie soorten)
- Basilemys
- Boremys
- Judithemys
- Neurankylus
- Plesiobaena
- Twee onbepaalde taxa
Zoogdieren
[bewerken | brontekst bewerken]Overblijfselen van de volgende zoogdieren zijn gevonden in de formatie:
- Multituberculata
- Cimexomys sp.
- Cimolodon spp.
- Cimolomys clarki
- Meniscoessus major
- Mesodma primaeva
- Naamloze multituberculaten
- Metatheria
- Alphadon halleyi
- Eodelphis browni
- E. cutleri
- Vijf soorten van Pediomys
- Turgidodon russelli
- T. praesagus
- Eutheria
- Cimolestes sp. (onzekere taxonomie)
- Gypsonictops lewisi
- Paranyctoides sternbergi
- Onbekende Theria: minimaal een soort
Vissen
[bewerken | brontekst bewerken]Overblijfselen van de volgende vissen zijn gevonden in de formatie:
- Chondrichthya
- Cretorectolobus olsoni (een tapijthaai)
- Eucrossorhinus microcuspidatus (een tapijthaai)
- Ischyrhiza mira (een sclerorhynchide)
- Meristodonoides montanensis (een haai)
- Myledaphus bipartitus (een rog)
- Protoplatyrhina renae (een gitaarvis)
- onbepaalde orectolobide
- Acipenseriformes (steuren)
- Acipenser albertensis
- Anchiacipenser acanthaspis
- Naamloze steur
- Naamloze lepelsteur
- Holostei
- Lepisosteus occidentalis (kaaimansnoek)
- Naamloze moddersnoek
- Ten minste twee andere Holostea
- Teleostei
- Belonostomus longirostris
- Cretophareodus (een osteoglossomorf)
- Coriops amnicolus
- Estesesox foxi
- Oldmanesox
- Paralbula (including Phyllodus)
- Paratarpon apogerontus (een elopomorf, als de tarpon)
- Ten minste acht andere Teleostei
Invertebraten
[bewerken | brontekst bewerken]Overblijfselen van de volgende ongewervelde dieren zijn gevonden in de formatie:
- Zoetwatertweekleppigen
- Fusconaia
- Lampsilis
- Sphaerium (twee soorten)
- Zoetwater slakken
- Campeloma (twee soorten)
- Elimia
- Goniobasis (drie soorten)
- Hydrobia
- Lioplacodes (twee soorten)
Flora
[bewerken | brontekst bewerken]Fossielen van plantenlichamen
[bewerken | brontekst bewerken]De volgende plantenfossielen zijn gevonden in de formatie:
- Diverse varens
- Equisetum (Equisetaceae)
- Gymnospermen
- Platyspiroxylon (Cupressaceae)
- Podocarpoxylon (Podocarpaceae)
- Elatocladus (Taxodiaceae)
- Sequoia (Taxodiaceae)
- Sequoiaxylon (Taxodiaceae)
- Taxodioxylon (Taxodiaceae)
- Ginkgo's
- Baiera
- Ginkgoites
- Angiospermen
- Artocarpus (Moraceae)
- Cercidiphyllum (Cercidiphyllaceae)
- Dombeyopsis (Sterculiaceae)
- Menispermites (Menispermaceae)
- Pistia (Araceae)
- Platanus (Platanaceae)
- Vitis (Vitaceae)
- Trapa (Trapaceae)
Palynomorpha
[bewerken | brontekst bewerken]Palynomorfen zijn microfossielen met een organische wand, zoals sporen, pollen en algen. De volgende palynomorfen zijn gevonden in de formatie:
- Onbekende producenten
- Ten minste acht soorten
- Fungi
- Ten minste vijfendertig taxa
- Chlorophyta (groene en blauwgroene algen)
- Ten minste twaalf soorten
- Pyrrhophyta (dinoflagellaten, een type van mariene algae)
- niet toegewezen cysten
- Bryophytes (mossen, levermossen en hauwmossen)
- Anthocerotophyta (hauwmossen)
- Ten minste vijf soorten
- Marchantiophyta (levermossen)
- Ten minste veertien soorten
- Bryophyta (mossen)
- Ten minste vijf soorten
- Anthocerotophyta (hauwmossen)
- Lycopodiophyta
- Lycopodiaceae (wolfsklauwen)
- Ten minste elf soorten
- Selaginellaceae (kleine wolfsklauwen)
- Ten minste zes soorten
- Isoetaceae (isoëtes)
- Ten minste één soort
- Lycopodiaceae (wolfsklauwen)
- Polypodiophyta
- Osmundaceae (koningsvarens)
- Ten minste zes soorten
- Schizaeaceae (klimvarens)
- Ten minste twintig soorten
- Gleicheniaceae (Gleichenia en allies; koraalvarens)
- Ten minste vijf soorten
- Cyatheaceae (Cyathea en allies)
- Ten minste vier soorten
- Dicksoniaceae (Dicksonia en allies)
- Ten minste drie soorten
- Polypodiaceae (varens)
- Ten minste vier soorten
- Matoniaceae
- Ten minste één soort
- Marsileaceae
- Ten minste één soort
- Osmundaceae (koningsvarens)
- Pinophyta (gymnospermen)
- Cycadaceae (cycaden)
- Ten minste drie soorten
- Caytoniaceae
- Ten minste één soort
- Pinaceae (dennen)
- Ten minste vier soorten
- Cupressaceae (cipressen)
- Ten minste drie soorten
- Podocarpaceae (Podocarpus en allies)
- Ten minste vier soorten
- Cheirolepidiaceae
- Ten minste twee soorten
- Ephedraceae (ephedra)
- Ten minste zes soorten
- Onbekende gymnospermen: Ten minste 3 soorten
- Cycadaceae (cycaden)
- Magnoliophyta (angiospermen)
- Magnoliopsida
- Buxaceae (buxus)
- Ten minste één soort
- Gunneraceae (mammoetblad)
- Ten minste één soort
- Salicaceae (wilgen, katoenhout, ratelpopulier)
- Ten minste één soort
- Droseraceae (zonnedauw)
- Ten minste één soort
- Olacaceae
- Ten minste twee soorten
- Loranthaceae
- Ten minste één soort
- Sapindaceae (zeepboom)
- Ten minste één soort
- Aceraceae (esdoorns)
- Ten minste één soort
- Proteaceae
- Ten minste negen soorten
- Compositae (composieten)
- Ten minste één soort
- Fagaceae (beuken, eiken, kastanjes)
- Ten minste twee soorten
- Betulaceae (berken, elzen)
- Ten minste één soort
- Ulmaceae (iepen)
- Ten minste één soort
- Chenopodiaceae
- Ten minste één soort
- Buxaceae (buxus)
- Liliopsida (eenzaadlobbigen)
- Liliaceae (lelies)
- Ten minste zes soorten
- Cyperaceae (zegge)
- Ten minste één soort
- Sparganiaceae (egelskop)
- mogelijk één soort
- Onbekende angiospermen: Ten minste achtentachtig soorten
- Liliaceae (lelies)
- Magnoliopsida
Tijdlijn van nieuwe taxa
[bewerken | brontekst bewerken]De volgende tijdlijn toont geldige taxa die voor het eerst zijn ontdekt in de Dinosaur Park Formation. Sommige soorten zijn mogelijk na hun oorspronkelijke beschrijving aan andere geslachten toegewezen.

- Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Dinosaur Park Formation op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar.