Anatomija pasa
Anatomija psa obuhvata anatomske studije vidljivih dijelova tijela pasa. Detalji o strukturi se izuzetno razlikuju od pasmine do pasmine, više nego kod bilo koje druge životinjske vrste, divlje ili pripitomljene,[1] jer su psi vrlo promjenjivi u visini i težini. Najmanji poznati odrasli pas je jorkširski terijer koji je imao visinu samo 6,3 cm do ramena, a 9,5 cm dužine duž glave i tijela. Težio je samo 113 g. Najveći poznati odrasli pas bio je engleski mastif koji je težio 155,6 kg i bio je dug 250 cm, od njuške do repa.[2] Najviši poznati odrasli pas je njemačka doga sa 106,7 cm do ramena.[3]

2. Njuška
3. Podbradak (vrat, koža vrata)
4. Rame
5. Lakat
6. Prednje noge
7. Sapi (krsta)
8. Noga (bedro i kuk)
9. Skočni zglob
10. Stražnje noge
11. Greben
12. Koljeno
13. Šape
14.Rep
Anatomija
[uredi | uredi izvor]Izvor:[4]
Mišići
[uredi | uredi izvor]Slijedi popis mišića psa, zajedno s njihovim polazištem, hvatištem, djelovanjem i inervacijom.
- Ekstrinzični mišići grudnog ekstremiteta i srodne strukture
Silazna površna grudnog koša: plazište od prve grudne kosti i hvatište na velikoj kvržicii humerusa. Obje privodi ekstremitetu i sprečava odmicanje ekstremiteta tokom nošenja tereta. Inerviraju ga kranijskii grudnii živci.
Poprečna površna grudne kosti: polazište na drugoj i trećoj grudnoj kosti i hvatište na velikoj kvržici humerusa. Također primiče ud i sprečava njegovo odmicanje tokom nošenja tereta. Inerviraju ga kranijalni grudni živci.
Duboki grudni koš: polazište je na trbušnoj strani grudne kosti a hvatište na maloj kvržici nadlaktične kosti. Djeluje na ekstenziju ramenog zgloba tokom nošenja tereta i savijanje ramena kada nema težine. Inerviraju ga stražnjii grudni živci.
Sternocephalicus: polazište sa sternuma i hvtište na sljepoočnoj kosti glave. Njegova je funkcija pomicanje glave i vrata s jedne na drugu stranu. Inervira ga pomoćni živac.
Sternohyoideus: polazište od grudne kosti i hvatište na bazihioidnoj kosti. Njegova je funkcija kaudalno pokretatanje jezika. Inerviraju ga trbušne grane vratnih kičmenih nerava.
Sternothyoideus: polazište od prve obalne hrskavice i hvatište na štitnjači. Njegova je funkcija i kaudalno pomicanje jezika. Inerviraju ga trbušne grane vratnih kičmenih živaca.
Omotransversarius: polazište na grebenu lopatice i hvatište na krilu atlasa. Njegova je funkcija pomicanje uda i bočno savijanje vrata. Inervira ga pomoćni živac.
Trapezius: polazište od supraspinoznog ligamenta, a hvatište na kičmi lopatice. Njegova je funkcija je podizanje i odmicnje prednjih ekstremiteta. Inervira ga pomoćni živac.
Rhomboideus: polazište od lopatičnog grebena potiljačne kosti, a hvatište na lopatici. Njegova je funkcija podizanje prednjih ekstremiteta. Inerviraju ga trbušne grane kičmenihpršljenova. . Latissimus dorsi: polazište na grudnoslabinskoj fasciji – hvatište na glavnom zadebljanju nadlaktične kosti. Njegova funkcija je savijanje ramenog zgloba. Inervira se torakodorzalnim živcem. Serratus ventralis: polazište od poprečnih nastavaka posljednjih pet vratnih pršljenova i hvatište na lopatici. Njegova je funkcija podrška trupa i pritiskivanje lopatice. Inerviraju ga trbušne grane vratnih kičmenih nerava.
- Unutrašnji mišići grudnog ekstremiteta
Deltoideus: polazište iz akromijalnog nastavka lopatice i hvatište na deltoidnom izraštaju zhadebljanju. Djeluje na savijanje ramena. Inervira ga pazušni živac.
Infraspinatus: polazište iz fossa infraspinatus, a hvatište i na velikoj kvržicii humerusa. Djeluje na istezanje i savijanje ramenog zgloba. Inervira se supralopatičnim živcem.
Teres minor: polazište od infraglenoidnog izraštaja na lopatici i hvatište se na malom teresnom izraštaju nadlaktične kosti. Djeluje na savijanje ramena i okretanje ruke bočno. Inervira ga pazušni živac.
Supraspinatus: polazište na supraspinoznoj jami i hvatište na većoj kvržicii humerusa. Djeluje na ekstenziju i stabiliziranje ramenog zgloba. Inervira ga supralopatični živac.
- Medijalni mišići lopatice i ramena
- '
Subscapularis: polazište od podlopatične jame – hvatište na velikoj kvržici humerusa. Djeluje medijalno, zakrećući prednju nogu i stabilizirajući zglob. Inervira ga sublopatični živac.
Teres major: polazište od lopatice i hvatište na glavnom izraštaju teresa nadlaktične . Djeluje na savijanje ramena i medijalnu rotaciju noge. Inervira ga pazušni nerv.
Coracobrachialis: polazište od korakoidnog nastavka lopatice – hvatište na grebenu donje kvržice nadlaktične kosti. Djeluje na primicanje, ekstenziju i stabilizaciju ramenog zgloba. Inervira se mišićno-kožnim živcem.
- Kaudalni grudni mišići
Tensor fasciae antebrachium: polazište od fascije koja pokriva latissimus dorsi – hvatište na olekranonskom nastavku. Djeluje na ekstenziju lakta. Inervira ga radijusni živac.
Triceps brachii: polazi od kaudalne granice lopatice – hvatište na zadebljanju olekranona. Djeluje na istezanje lakta i savijanje ramena. Inervira ga radijumski živac.
Anconeus: polazište od nadlaktične kosti i hvatište na proksimalnom kraju ulne. Djeluje na ekstenziju lakta. Inervira ga radijusni živac.
- Kranijski mišići prednje noge
Biceps brachia: polazi od supraglenoidne kvržice – hvatište na ulninom i radijusovom izrašrtaju. Djeluje na savijanje lakta i istezanje ramena. Inervira se mišićno-kožnim živcem.
Brachialis: polazi od bočne površine nadlaktične kosti – hvatište na ulninom i radijusnom izraštaju. Djeluje na savijanje lakta. Inervira se mišićno-kožnim živcem.
- Kranijski i bočni antebrahijski mišići
Extensor carpi radialis: potječe na suprakondilarnom grebenu i umeci na metakarpalima. Djeluje na ekstenzijunerv karpusa. Inervira ga radijalni nervom.
Zajedničkii ekstenzor prstiju: polazište na bočnom epikondilu nadlaktične kosti – hvatište na distalnim falangama. Djeluje na ekstenziju šake i zglobova 3., 4. i 5. Prsta. Inervira ga radijusni živac.
Extensor carpi ulnaris: polazi od bočnog epikondila nadlaktične kosti – hvatište na metakarpusnoj 5. i pomoćnoj karpusnoj kosti. Djeluje na odmicanje i ekstenziju karpusnog zgloba. Inervira ga radijusni nerv.
Supinator: polazište na bočnom epikondilu nadlaktične kosti, a hvatište na radijusu. Djeluje na bočno okretanje podlaktice. Inervira ga radijusni nerv.
Abductor pollicis longus: polazište sa ulne – hvatište na 1. metakarpusnoj kosti. Djeluje tako da odmiče prst, uz ekstenziju karpusnih zglobova. Inervira ga radijusni nerv.
- Kaudalni i medijalni mišići podlaktice
Pronator teres: polazi na medijalnom epikondilu nadlaktične kosti – hvatište na medijalnoj granici rfadijusa. Djeluje medijalno zakrećući podlakticu i savijajući lakat. Inervira se medijalnim nervom.
Flexor carpi radialis: polazi od medijalnog epikondila nadlaktične kosti – hvatište na dlanskoj strani 2. i 3. metakarpusne kosti. Djeluje na savijanje karpusa. Inervira se srednjim živcem.
Površinski fleksor prstiju: polazi od medijalnog epikondila nadlaktične kosti i hvata na dlanskoj površini srednjih falangi. Djeluje na savijanje karpusnih, metakarpofalangenih i proksimalnih interfalangnih zglobova prstiju. Inervira se srednjim nervom.
Flexor carpi ulnaris: polazište od olekranona, sa hvatištem na pomoćnoj karpusnoj kosti. Djeluje na savijanje karpusa. Inervira ga ulnini nerv. Duboki fleksor prstiju: polazi od medijalnog epikondila nadlaktične kosti i hvata se za dlansku površinu distalnih falangi. Djeluje na savijanje karpusa, metakarpofalangenih zglobova te proksimalnih i distalnih interfalangenih zglobova prstiju. Inervira se srednjim nervom.
Pronator quadratus: polazište od površina radijusa i ulne. Djeluje na pronaciju šape. Inervira se srednjim nervom.
- Kaudalni mišići bedra
Biceps femoris: polazi od ishiatske zadebljale pločice i hvata se na ligamentu patele. Djeluje na ekstenziju kuka, ukočenost i skočni zglob. Inervira ga ishijadični nerv.
Semitendinosus: polazište od ishiatske zadebljale pločice – hvatište na tibiji. Djeluje na istezanje kuka, savijanje koljena i ekstenziju skočnog zgloba. Inervira ga ishijadični nerv.
Semimembranosus: polazi od ishiatske izrasline, sa hvatištem na femuru i tibiji. [Djeluje na ekstenziju kuka i ukočenost. Inervira ga išijasni nerv.
- Medijalni mišići bedra
Sartorius: potječe iz ilijuma i umetaka na pateli i tibiji. Djeluje na savijanje kuka i na savijanje i produžavanje koljena. Inervira ga bedreni živac.
Gracilis: polazište iz karlične simfize – hvatište na kranijalnoj granici tibije. Djeluje na primicanje uda, savijanje koljena i ekstenziju ekstenziju kuka i skočnog zgloba. Inervira ga zatezni živac.
Pektineus: polazište iz iliopubične eminencije, a hvatište na repnom dijelu butne kosti. Djeluje na adukciju uda. Inervira ga zatezni živac.
Primicač: polazi iz karlične simfize –hv atište na bočnoj strani butne kosti. Djeluje na adukciju uda i ekstenziju kuka. Inervira ga zatezni živac.
- Lateralni mišići karlice
Tensor fasciae latae: polazište od koljenskog zadebljanja ilijuma i hvatište na bočnoj fasciji femura. Djeluje na savijanje kuka i ekstenziju koljena. Inervira ga kranijalni gluteusni živac.
Površinski gluteus: polazište na bočnoj granici krstačne kosti – hvatište na 3. trohanteru. Djeluje naekstenziju kuka i odmicanje uda. Inervira ga repni gluteusni živac.
Srednji gluteus: polazište od ilijuma i hvatište na velikom trohanteru. Djeluje tako na odmicsnje kuka i medijalno rotira karlični ud. Inervira ga kranijalni gluteusni živac.
Duboki gluteus: polazi od ishiatskog dijela kičme – hvatište na velikom trohanteru. Djeluje na ekstenziju kuka i medijalnu rotaciju karličnog uda. Inervira ga kranijalni gluteusni živac.
- Kaudalni mišići kuka
Internal obturator: polazište od karlične simfize i hvatište na trohanternoj jami femura. Djeluje na bočnu rotaciju karličnog uda. Inervira ga išijasni živac.
Gemelli: polazište sa bočne površine ishijuma i hvatište na trohanternoj jami. Djeluje na bočnu roraciju karličnog uda. Inervira ga išijasni živac.
Quadratus femoris: polazište od ishiuma i hvatište na intertrohanternom grebenu. Djeluje na rkstenziju kuka i bočno rotiranje karličnog uda.
Vanjski opturator: polazište od pubisa i ishijuma i hvatište na trohanternoj jami. Djeluje na bočnu rotaciju karličnog uda. Inervira ga zatezni živac.
- Kranijalni mišići bedra
Quadriceps femoris': polazišteod femura i ilijuma i hvatište na zadebljanju tibije. Djeluje na ekstenziju koljena i savijanje kuka. Inervira ga bedreni živac.
Ilipsoas: polazište na ilijumu i hvatištena malom trohanteru. Djeluje na savijanje kuka. Inervira ga bedreni živac.
- Kraniolateralni mišići noge
Kranijalni tibialis: polazište tibije i hvatište na tab anskoj površini 1. i 2. metatarzusne kosti. Djeluje na savijanje tarzusa i bočno rotiranje šape. Inervira ga peroneusni živac.
Dugi ekstenzor prstiju: polazište iz ekstenzorske jame femura i hvatište u ekstenzorskim nastavcima distalnih falangi. Djeluje na ekstenziju prstiju i savijanje stopala. Inervira ga peroneusni živac.
Peroneus longus: polazište na tibiji i fibuli, a hvatištena 4. tarzusnoj kosti i tabanskoj površini metatarzusa. Djeluje na savijanje tarzusa i medijalo rotiranje šape. Inervira ga peroneusni živac.
- Kaudalni mišići noge
Gastrocnemius: polazište od suprakondilusnih zadebljanj bedrene kosti i hvatište na kalkaneusnoj izbočini. Djeluje na ekstenziju stopala i savijanje koljena. Inervira ga tibijski živac.
Površinski fleksor prstiju: polazište od bočne suprakondilusne izbočine femura, a hvatište na kalkanusnim izbočinama i osnovama srednjih falangi. Djeluje na savijanje potkoljenice i ekstenziju stopala. Inervira ga tibijski živac.
Duboki fleksor prstiju: polazište na fibuli i hvatište na tabanskoj površini distalnih falangi. Djeluje na savijanje prstiju i ekstenziju tarzusa. Inervira ga tibijski živac.
Popliteus: polazište od bočnog kondilusa bedrene kosti – hvatište na tibiji. Djeluje na medijalno zrotiranje nogu. Inervira ga tibijski živac.
Skelet
[uredi | uredi izvor]- Kosti i njihove značajne tačke za vezanje mišića:
Lopatica:' Kičma lopatice, supraglenoidna kvržica, glenoidna šupljina, akromionski nastavak, supraspinozna jama, infraspinozna jama, vrat, korakoid, nastavak, podlopatična jama;
Humerus: Glava humerusa, veelika kvržica, maala kvržica, međukvržični žlijeb, deltopektoralni greben, deltoidna izlaslina, tijelo humerusa, epikondili (medijalni i bočni), humerusni kondil (trohleja i kapitulum, radijalne i olekranonske jame)
- Ulna i radijus:
Olekranonski nastavak, trohleusni urez, ankoneusni nastavak, koronoidni nastyvci (medijalni i bočni), tijelo ulne, glava radijusa, tijelo radijusa, distalna trohlea, stiloidni nastavak (medijalni i bočni), međukostni prostor;
Korijen šake Kosti šake (radijusne i ulnine), Pomoćnene karpusne kosti, 1. 2. 3. I 4. kost korijena šake, falange, proksimalna osnova, tijelo, glava, ungualni greben, negualni nastavak, ekstenzorski nastavak proces, karpometakarpusni zglobovi, metakarpofalangni zglobovi, proksimalni međufalangni zglobovi, interfalangni zglobovi
- Glava, ligament glave, vrat, veliki trohanter, mali trohanter, trohanterska jama, acetabulumska jama (na bokovima), distalna butna kost, trohlea (i grebeni), kondili (medijalni / bočni), epikondili (medijalni / bočni), međukondilusna jama, ekstenzorska jama, infračašična masna podloga, fabele (medijalno / bočno)
(koljenska kapa)
Kondili tibije (medijalni / bočni), međukondilskee eminencije, ekstenzorski urez (bočni), tibijska izraslina (kranijalna), tibijska kohlea, meedijalni maleolus, bočni maleolus, glava fibule
- Talus, kalkaneus, trohanterski grebenovi, centralna kost stopala, 1. 2. i 3. kost stopala
- Tijelo, peteljke, laminae, spinozni nastavak, poprečni nastavak (krila), zglobni nastavak, kičmeni otvor, međupršljenski otvori, atlas (pršljen C-1),
epistrofeus (C2), jame, ventralna ploča (na pršljenu C-6);

2. Gornja vilica
3. Donja vilica
4. Atlas
5. Epistrofeus
6. Lopatica
7. Lopatični trnoliki nastavak
8. Humerus
9. Radijus
10. Ulna
11. Falange 12. Kosti korijena šake
13. Kosti šake
14. Sternum
15. Hrskavičavi dio rebra
16. Rebra
17. Falange
18. Metatarzus
19. kosti stopala
20. Kalkaneus
21. Fibula
22. Tibija
23. Patela
24. Femur
25. Ishium
26. Pelvis
- Sastav
-
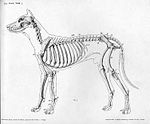 Skelet psa, bočno
Skelet psa, bočno -
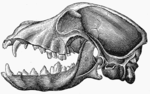 Pasja lobana sa otvorenim vilicama, bočno
Pasja lobana sa otvorenim vilicama, bočno -
 Bočni izgled lobanjae psa
Bočni izgled lobanjae psa -
 Čeoni izgled lobanje
Čeoni izgled lobanje -
 Pasji zubi
Pasji zubi





- Lobanja
U 1986. Godine, studija morfologije lobanje otkrila je da se kod domaćeg psa morfološki razlikuje od svih ostalih kanida, osim od vučje. Razlika u veličini i udjelu među nekim pasminama jednako je velika kao i kod bilo kojeg divljeg roda, ali svi su psi očito pripadnici iste vrste.[5] U 2010., istraživanje oblika lobanje psa u odnosu na postojeće mesojede predložilo je da "najveća udaljenost oblika između pasmina pasa očito premašuje maksimalnu divergenciju između vrsta mesojedaa. Štaviše, domaći psi zauzimaju čitav niz novih oblika izvan domena divljih mesojeda".[6]
Domaći pas, u poređenju s vukom, pokazuje najveće razlike u veličini i obliku lobanje (Evans 1979) koje se kreću od 7 do 28 cm dužine (McGreevy 2004). Vukovi su dolihokefalni (dugoglavi), ali ne tako ekstremni kao neke pasmine pasa, kao što su hrtovi i ruski vučjaci (McGreevy 2004). Pasja brahikefalija (kratkoglavost) nalazi se samo kod domaćih pasa i srodna je pedomorfozi (Goodwin 1997). Štenad se rađa s kratkim njuškama, a duža lobanja dolihocefalnih pasa pojavljuje se u kasnijem razvoju (Coppinger 1995). Ostale razlike u obliku glave između brahikefalnih i dolihokefalnih pasa uključuju promjene kraniofacijalnog ugla (između bazilarne osi i tvrdog nepca) (Regodón 1993), morfologiju sljepoočnomandibulskog zgloba (Dickie 2001) i radiografsku anatomiju kribriformne ploče (Schwarz 2000).[7]
Jedno istraživanje pokazalo je da je relativno smanjenje dužine lobanje psa, u odnosu na njenu širinu (Kefalični indeks) značajno povezano i sa položajem i sa uglom mozga u lobanji. Tako je bilo bez obzira na veličinu mozga ili tjelesnu težinu psa.[8]

| Kanidni | Karnivorski | Očnjak |
|---|---|---|
| Vuk | 131,6 | 127,3 |
| Azijski divlji pas | 130,7 | 132,0 |
| Afrički divlji pas | 127,7 | 131,1 |
| Grenlandski pas (pripitomljeni) | 117,4 | 114,3 |
| Kojot | 107,2 | 98,9 |
| Bočnoprugasti šakal | 93,0 | 87,5 |
| Zlatni šakal | 89,6 | 87,7 |
| Crnoleđi šakal | 80,6 | 78,3 |
Respiratorni sistem
[uredi | uredi izvor]
Respiratorni sistem je skup organa odgovornih za unos kisika i izbacivanje ugljik-dioksida.
Kako psi imaju malo znojnih žlijezda u koži, dišni sistem također ima važnu ulogu u tjelesnoj termoregulaciji.[10]
Psi su sisari s dva velika plućna krila koja su dalje podijeljena u režnjeve. Spužvastog su izgleda zbog prisustva sistema nježnih grana bronhiola u svakom od njih, koje završavaju zatvorenim komorama s tankim zidovimama (tačke izmjene plinova) zvanim alveole .
Prisustvo išićne strukture, dijafragme, ekskluzivne za sisare, dijeli peritoneumsku šupljinu od pleurne šupljine, osim što pomaže plućima tokom udisanja.
Probavni sistem
[uredi | uredi izvor]Organi koji čine pseći probavni sistem su:[11]
-
 Psećji cekum.
Psećji cekum. -
 Probavni sistem psa
Probavni sistem psa -
 Psećji stomak.
Psećji stomak. -
 Psećji stomak (unutrašnjost)
Psećji stomak (unutrašnjost) -
 Psećji jezik, fiksiran u formalinu
Psećji jezik, fiksiran u formalinu -
 Psećji ileum.
Psećji ileum. -
 Vaskularna struktura psećje jetre.
Vaskularna struktura psećje jetre.



.jpg)



Reproduktivni organi i razmnožavanje
[uredi | uredi izvor]Muški spolni organi
[uredi | uredi izvor]
Genitalije psa su spolne žlijezde – sjemenici s epididimisom trajno smješteni u mošnjama, gdje se obično spuštaju do dva mjeseca nakon rođenja. Epididimusni kanal nastavlja se vas deferens, koji se otvara u mokraćovod na sjemenom grebenu. Pas ima jednu dodatnu spolnu žlijezdu, relativno veliku prostatu, koja je približno veličine lješnika do kestena. Mokraćovod se otvara na vrhu penisnog glavića. Penis psa proteže se od sjedne kosti sa dva korijena i tvore ga aksilarna tijela. Glans je izdužen i također ima aksilarno tijelo, koje na svom stražnjem dijelu čini bulbus glandis. Vrh penisa također je ojačan s penisnom kosti. Cjelokupni penis doseže do pupka i obavijen je u kožicu, odakle se proteže tokom uzbuđenja.
Ženski spolni organi
[uredi | uredi izvor]
Ženske spolne žlijezde, jajnici, nalaze se u trbušnoj šupljini, iza bubrega na razini 3-4. slabinskog pršljena i veličine su 1–2 cm. Ima oblik tanke cijevi dužine 4–7 cm, koja se otvaraj na vrhu odgovarajućeg roga maternice. Ženka ima dvorogu Y-oblikovanu maternicu . Kujina vagina je dugačka i završava na vaginalnskm ulazu, koji je kod novorođenih kuja zaštićen djevičanskim himenom. Vaginu na ušću mokraćovod prati koso vaginsko perdvorje, koje je okruženo aksilarnim tijelom "bulbusnih predvorja". U sluznici vagine nalaze se glandulae vestibulares minores . Vanjske genitalije čine vulva, koja se sastoji od dvije labije. Usnama upravljaju kratki mišići, zahvaljujući kojima je vulva pokretna i tokom parenja, tako da se kuja može aktivno približiti tamo gdje osjeća penis mužjaka.
U domaćih pasa spolna zrelost (pubertet) počinje se javljati u dobi od šest do dvanaest mjeseci i kod mužjaka i kod ženki, iako u nekih velikih pasmina može se odgoditi do druge godine. Adolescencija većine pasa traje između dvanaest i petnaest mjeseci starosti, od tada su već odrasliji od štenadi. Kao i kod drugih udomaćenih vrsta, pripitomljavanje je potaknulo veći libido i raniji i češći ciklus parenja pasa nego kod njihovih divljih predaka. Psi ostaju reproduktivno aktivni i do starosti.
Većina pasa svoj prvi spolni žar ima u dobi od šest do dvanaest mjeseci, iako ga neke pasmine velikih nemaju do druge godine. Ženke ulaze u libido dva puta godišnje, tokom kojeg se njihova tijela pripremaju za trudnoću, a na vrhuncu ulaze u estrus, razdoblje tokom kojeg su etološki i fizički prihvatljive za kopulaciju. Budući da jajašca opstaju i mogu biti oplođena I sedmicu dana nakon ovulacije, moguće je da se ženka pari s više mužjaka.
Kuje rađaju 56–72 dana nakon oplodnje, s prosječno 63 dana, iako dužina trudnoće može varirati. Prosječno leglo sastoji se od oko šestoro štenadi , iako se taj broj može uveliko razlikovati ovisno o pasmini pasa. Psi “igračke” rađaju između jednog i četiri šteneta po leglu, dok veće pasmine mogu u prosjeku dati dvanaest štenaca po leglu. Neke pasmine stekle su selektivnim uzgojem svojstva koji ometaju reprodukciju. Naprimjer, mužjaci francuskih buldoga ne mogu zajahati ženku. U velikoj većini slučajeva, ženke ove pasmine moraju se umjetno oploditi kako bi se mogle razmnožavati.
Fizička svojstva
[uredi | uredi izvor]
Kao i većina grabežljivih sisara, pas ima snažne mišiće, kardiovaskularni sistem koji podržava sprint i izdržljivost, te zube za hvatanje, držanje i kidanje.
Mišići psa pružaju mogućnost skakanja i odskakanja. Noge ih mogu brzo gurnuti naprijed, skačući po potrebi kako bi progonili i svladali plijen. Imaju mala, uska stopala, hodajući na prstima (tako da imaju digitigradni stav i kretanje). Stražnje noge su im prilično krute i čvrste. Prednje su labave i fleksibilne, a samo ih mišići pričvršćuju za trup.
Veličina njuške psa ovisit će o pasmini. Psi sa srednjim njuškama, kao što je njemački ovčar, nazivaju se mezocefalni, a psi s gurnutom njuškom, poput Mopsa, brahikefalni. Današnje pasmine igračka imaju kosture koji sazrijevaju za samo nekoliko mjeseci, dok divovskim pasminama, poput mastif treba 16 do 18 mjeseci da kostur sazri. Patuljavost uticala je na proporcije kostura nekih pasmina, kao u psa rase baset
Svi psi (i svi današnji predstavnici porfodice Canidae) imaju ligament koji povezuje trnoliki nastavak prvog grudnog pršljena sa stražnjom stranom osi kosti (druga vratna kost), koja podupire težinu glave, bez aktivnog naprezanja mišića, čime se štedi energija.[12] Ovaj je ligament po funkciji analogan (ali različit u točnim strukturnim detaljima) onom nuhalni ligament pronađen u ungulate s.[12] Ovaj ligament omogućuje psima da nose glavu dok trče na velike daljine, naprimjer dok prate miris staze nosom do zemlje, bez trošenja puno energije.
Psi imaju odvojene kosti ramena (nedostaje vratna kost ljudskog kostura), što omogućava veću dužinu koraka za trčanje i skakanje. Hodaju na četiri prsta, sprijeda i straga, a na prednjim i stražnjim nogama imaju zakržljale čaponjke. Kad pas osim uobičajenog stanja, otraga ima i dodatne, kaže se da je pas "dvostruko čaponjast". Veličina njuške psa ovisit o pasmini. Psi sa srednjim njuškama, kao što je njemački ovčar, nazivaju se mezokefalni, a psi s gurnutom njuškom, poput mopsa, brahikefalni. Današnje pasmine igračkih pasa imaju kosture koji sazrijevaju za samo nekoliko mjeseci, dok džinovskim pasminama, poput mastifa, treba 16 do 18 mjeseci da kostur sazri. Patuljavost uticala je na proporcije kostura nekih pasmina, kao u pas baset.
Veličina
[uredi | uredi izvor]
Psi se jako razlikuju u visini i težini. Najmanji poznati odrasli pas bio je jorkširski terijer, koji je bio visok samo 6,3 cm do ramena, a 9,5 cm dug, a težio je samo 113 g. Najveći poznati odrasli pas bio je engleski mastif koji je težio 155,6 kg i bio je dug 250, od njuške do repa.[2] Najviši poznati odrasli pas je njemačka dogam visine 106,7 cm, do ramena.
U 2007. jedna studija je identifikovala gen koji je predložen kao odgovoran za veličinu. Otkrila je regulacijsku skvencu, koja, pored gena za Insulinoliki faktor rasta 1 (IGF1), zajedno sa genom i regulatornom sekvencom ima "glavni doprinosi veličini tijela kod svih malih pasa". Dvije su varijante ovog gena pronađene kod velikih pasa, što čini složeniji razlog velikih dimenzija pasmine. Istraživači su zaključili da upute ovog gena da psi budu mali moraju biti stare najmanje 12.000 godina i da ga nema u vukova.[13] Prema drugom istraživanju, „psi za krilo“ (mali psi) su među najranijim postojećim psima.[14]
Dlaka
[uredi | uredi izvor]
Domaći psi često imajuuju ostatke kontraobojenja, uobičajenog prirodnog oblika kamuflaže. Općenita teorija kontrasjenčenja je da će se životinja koja je osvijetljena odozgo na gornjoj polovini činiti svjetlijom, a na donjoj tamnijom, gdje će obično biti u sopstvenoj sjenci.[15][16] Ovo je obrazac na koji grabljivice mogu naučiti paziti na okruženje. Životinja u kontrasjeni imat će tamne boje na gornjim površinama, a svijetle boje ispod.[15] To smanjuje opću vidljivost životinje. Podsjetnik na ovaj obrazac je da će mnoge pasmine povremeno imati "biljeg", prugu ili "zvijezdu" bijelog krzna na prsima ili donjoj strani.[16]
Studija je otkrila da se genetička osnova koja objašnjava boje dlake kod konja i mačaka nije vrijedila za pse.[17] U projektu uzeti su uzorci od 38 različitih pasmina/rasa, kako bi se pronašao gen (beta defenzinski gen) odgovoran za boju dlake pasa. Jedna verzija proizvodi žute, a mutacija crne. Sve boje dlake za pse su modifikacije crne ili žute.[18] Naprimjer, bijelo u bijelom. što ima minijaturni šnaucer, je krem boje, a ne albino (genotip e/e na lokusu MC1R).
Današnje pasmine pasa pokazuju raznolik niz krznenih omotača, uključujući pse bez krzna, poput meksičkog bezdlakog psa. Pasje krzno razlikuje se u teksturi, boji i oznakama, a razvijen je i specijalizirani rječnik koji opisuje svako posebno.[19]
Rep
[uredi | uredi izvor]Postoji mnogo različitih oblika psećih repova: ravni, ravno prema gore, srpasti, uvijen i plutasti vijak. U nekih pasmina rep se obično skraćuje, kako bi se izbjegle ozljede (posebno za lovačke pse).[20] It can happen that some puppies are born with a short tail or no tail in some breeds.[21] Psi imaju ljubičastu ili nadrepnu žlijezdu na leđnoj (gornjoj) površini repa.
Šapa
[uredi | uredi izvor]Psi mogu dugo stajati, šetati i trčati po snijegu i ledu. Kad je pseća šapa izložena hladnoći, gubitak topline sprečava se prilagodbom krvnog sistema koji recirkulira toplinu natrag u tijelo. Donosi krv s površine kože i zadržava toplu krv na površini jastučića.[22]
Zaperak
[uredi | uredi izvor]Postoje neke rasprave o tome pomaže li zaperak (ili čaporak) psima da dobiju snagu pri trčanju, jer kod nekih pasa uspostavlja kontakt dok trče, a nokat na krznu često se istroši na isti način kao nokti na drugim nožnim prstima, od dodira sa zemljom. Međutim, kod mnogih pasačaporak nikada ne uspostavlja kontakt sa zemljom. U tom se slučaju nokat se nikada ne troši i tada se često obrezuje kako bi se zadržao na sigurnoj dužini.
Zaperci nisu mrtvi dodaci. Mogu se koristiti za lagano prihvatanje kostiju i drugih predmeta koje psi drže šapama. Međutim, kod nekih pasa možda se čini da ove kandže uopće nisu povezane s nogom, osim preklopkom kože. U takvih pasa te kandže nemaju svrhu hvatanja jer se lahko može saviti ili okrenuti.
Također se vodi rasprava o tome treba li ih hirurški ukloniti. Argument za uklanjanje navodi da su slab prst, jedva pričvršćen za nogu, tako da mogu djelomično otkinuti ili lahko zapeti za nešto i slomiti se, što može biti izuzetno bolno i sklono infekcijama. Drugi kažu da je bol prilikom uklanjanja čaporka daleko veća od bilo kojeg drugog rizika. Iz tog je razloga uklanjanje kandži i čaporka nezakonito u mnogim zemljama. Postoji, možda, iznimka za lovačke pse koji ponekad mogu pokidati ogrlicu dok trče u zarasloj vegetaciji. Ako se želi ukloniti čaporak, to bi trebalo biti učinjeno kada je pas štene, ponekad čak u dobi od 3 dana; iako se po potrebi može izvoditi i starijim psima (tada je operacija možda teža). Operacija je prilično jednostavna i može se izvesti samo uz primjenu lokalnih anestetika, ako prst nije dobro povezan s nogom. Nažalost, mnogi psi ne mogu odoljeti lizanju bolnih šapa nakon operacije, pa vlasnici moraju biti oprezni.
Osim toga, za one pse čiji čaporci kontaktiraju tlo dok trče, moguće je da bi uklanjanje moglo biti nedostatak za postizanje brzine psa u trčanju i promjeni smjera, posebno u sportskim prilikama kao što je agilnost pasa.
Čula
[uredi | uredi izvor]Vid
[uredi | uredi izvor]
Kao i većina sisara, i psi imaju samo dvije vrste čepićastih fotoreceptora, što ih čini dihromatima.[23][24][25][26] Ti čepići maksimalno su osjetljivi na talasnim dužinama između 429 nm i 555 nm. Studije ponašanja pokazale su da se pseći vizualni svijet sastoji od žutih, plavih i sivih nijansi,[26] ali imaju poteškoća u razlikovanju crvene i zelene, što njihov vid u boji čini ekvivalentnim crveno-zelenom sljepilu za boje kod ljudi (deuteranopija). Kad čovjek percipira objekt kao "crven", taj se psu čini "žut", a ljudska percepcija "zelenog" izgleda kao "bijela", sjena sive boje. Ova bijela regija (neutralna tačka) javlja se na oko 480 nm, dijelu spektra koji ljudima izgleda plavo-zeleno. Za pse se talasne dužine koje su veče od neutralne tačke ne mogu međusobno razlikovati i sve se pojavljuju kao žute.[26]
Psi koriste boju umjesto svjetline, kako bi razlikovali svijetlu ili tamnoplavu / žutu.[27][28][29] Manje su osjetljivi na razlike u sivim nijansama od ljudi i također mogu otkriti svjetlinu otprilike upola tačnije od ljudi.
Pasji vizualni sistem evoluirao je u pravcu prilagođavanja što uspješnijem lovu.[23] Iako je oštrina vida pasa loša (procjenjuje se da je pudlica prevedena u ocjenu Snellenove stope od 20/75, njihova vizualna diskriminacija je za pokretne predmete vrlo velika. Pokazalo se da psi mogu diskriminirati ljude (npr. identificirati skrbnika) u rasponu između 800 i 900 m; međutim, ovaj raspon smanjuje se na 500–600 m ako objekt miruje.
Psi mogu otkriti promjenu kretanja koja postoji u jednom dioptrijskom prostoru unutar svog oka. Za usporedbu, ljudima je potrebna promjena između 10 i 20 dioptrija da bi se otkrilo kretanje.[30]
Kao večernji lovci, psi se često oslanjaju na vid u situacijama pri slabom osvjetljenju: imaju vrlo velike zjenice, veliku gustoću štapića u fovea centralis, povećanu brzina treperenja i tapetum lucidum.[23] Tapetum je reflektirajuća površina iza mrežnjače, koja odbija svjetlost dajući fotoreceptorima drugu priliku za hvatanje fotona. Također postoji veza između veličine tijela i ukupnog promjera oka. Između različitih rasa pasa može biti raspon od 9,5 i 11,6 mm. Ova varijacija od 20% može biti značajna i pridružena je kao prilagodba vrhunskom noćnom vidu. Oči različitih pasmina pasa imaju različite oblike, dimenzije i konfiguracije mrežnjače.[31] Mnoge dugonose pasmine imaju "vizualnu crtu" – široku fovealnu regiju koja prolazi širinom mrežnjače i pruža im vrlo široko polje izvrsnog vida. Neke pasmine/rase sa dugom njuškom, posebno gonići, imaju vidno polje do 270° (u usporedbi s 180° za ljude). Pasmine kratke njuške, s druge strane, imaju area centralis: središnji dio s gustim nervnim završecima, do tri puta većom od vidnih traka, što im daje detaljniji vid mnogo sličniji ljudskom. Neke širokoglave pasmine s kratkim njuškom imaju vidno polje slično onom kod ljudi.

Većina pasmina ima dobar vid, ali neke pokazuju genetičku predispoziciju za miopiju – poput rotvajler a, za kojeg je utvrđeno da je svaki drugi kratkovidan.[23] Psi također imaju veće odstupanje očne osi od ljudi, što im omogućava da zjenice rotiraju dalje u bilo kojem smjeru. Divergencija očne osi pasa kreće se od 12 – 25°, ovisno o pasmini.[30]
Eksperimentiranjem je dokazano da psi mogu razlikovati složene vizualne slike, poput kocke ili prizme. Psi također pokazuju privlačnost prema statičnim vizualnim slikama, kao što je siluete psa na ekranu, vlastitih odraza ili video zapisa pasa; međutim, njihov interes naglo opada kada ne mogu uspostaviti socijalni kontakt sa slikom.
Sluh
[uredi | uredi izvor]
Raspon frekvencija psećeg sluha je između 16 i 40 Hz (u poređenju sa 20–70 Hz za ljude) i do 45–60 kHz (u poređenju sa 13–20 kHz za ljude), što znači da psi mogu detektirati zvukove daleko iznad gornje granice ljudskog slušnog spektra.[25][32][33][34]
Psi imaju pokretljivost ušiju, koja im omogućava da brzo utvrde tačnu lokaciju zvuka. Osamnaest ili više mišića može nagnuti, rotirati, podići ili spustiti pseće uho. Pas može prepoznati lokaciju zvuka mnogo brže nego što to može čovjek, kao i čuti zvukove na četverostruko većoj udaljenosti.[35]
Miris
[uredi | uredi izvor]
Dok ljudskim mozgom dominira veliki vizuelni korteks, psećim dominira veliki olfaktorni korteks.[23] Psi imaju otprilike četrdeset puta više receptori osjetljivi na miris od ljudi, u rasponu od oko 125 do gotovo 300 miliona kod nekih pasmina, poput krvoločnih pasa.[23] Smatra se da ovo njihov njuh čini do 40 puta osjetljivijim od ljudskog. Ovi receptori su rašireni na površini veličine džepne maramice (u usporedbi s 5 miliona na području veličine poštanske marke za ljude).[36][37] Njuh pasa takođe uključuje upotrebu vomeronosnog organa, koji se prvenstveno koristi za socijalne interakcije.
Pas ima pokretne nosnice koje mu pomažu u određivanju smjera mirisa. Za razliku od ljudi, ne treba napuniti pluća jer neprestano unosi miris u nos u naletima od 3-7 njušenja. Nos psa ima koštanu strukturu koju ljudi nemaju, što omogućava da zrak koji je njuškao prelazi preko koštane pregrade i na njega se lijepe mnogi molekule mirisne tvati. Zrak iznad ove police ne ispire se kad pas normalno diše, pa se molekule te molekule akumuliraju u nosnim komorama, a miris se intenzivno gradi, omogućavajući psu da detektira i najslabije mirise.
Jedno istraživanje sposobnosti pasa za učenje u usporedbi s vukovima pokazalo je da psi imaju bolji njuh od vukova kada lociraju skrivenu hranu, ali još uvijek nema eksperimentalnih podataka koji podupiru ovo gledište.[38]
Vlažni nos ili rinarij je neophodan za određivanje smjera zračne struje koja sadrži miris. Receptori hladnoće u koži osjetljivi su na njeno hlađenje isparavanjem vlage zračnim strujama.[39]
Okus
[uredi | uredi izvor]Psi imaju oko 1.700 okusnih pupoljaka, u poređenju sa ljudima sa oko 9.000. Pupoljci za slatko reagiraju na hemikaliju zvanu furaneol koja se nalazi u mnogim plodovima i paradajzu. Čini se da psi vole ovaj okus i vjerovatno je evoluirao jer u prirodnom okruženju psi često dopunjavaju prehranu malim životinjama, onim voćem koje je dostupno. Zbog odbojnosti pasa prema gorkom okusu, razni sprejevi i gelovi dizajnirani su kako bi ih spriječili grizu namještaj ili druge predmete. Psi također imaju okusne pupoljke koji su prilagođeni za vodu, što je nešto što dijele s drugim zvijerima, ali se kod ljudi ne nalazi. Ovaj osjećaj okusa nalazi se na vrhu psećeg jezika, koji je dio koji uvija ločuči vodu. Ovo područje reagira na vodu u svako doba, ali kada pas jede slanu ili slatku hranu, osjetljivost na okus vode se povećava. Smatra se ta se sposobnost razlikovanja okusa vode razvila kao način da tijelo održi ravnotežu unutrašnjih tekućina, nakon što životinja pojede nešto zbog čega će se izlučiti više mokraće ili će joj trebati više vode za adekvatnu obradu. Svakako se čini da kada su ovi posebni pupoljci za vodu aktivni, psi dobijaju dodatno zadovoljstvo od pijenja vode i da će popiti obilne količine.[40]
Dodir
[uredi | uredi izvor]
Glavna razlika između čula dodira čovjeka i psa je prisustvo specijaliziranih brkova. poznatih kao vibrisi. Vibrisi su prisutne iznad pasjih očiju, ispod vilica i na njušci. TO su sofisticirani organi za osjet. Krućei su i ugrađeni su mnogo dublje u kožu od ostalih dlačica i u osnovi imaju veći broj receptorskih ćelija. Mogu otkriti zračne struje, suptilne vibracije i predmete u mraku. Pružaju sistem ranog upozoravanja na predmete koji mogu udariti lice ili oči, a vjerovatno pomažu usmjeriti hranu i predmete prema ustima.[41]
Magnetna osjetljivost
[uredi | uredi izvor]Psi mogu radije vršiti nuždu kada su sklonjeni na uzici i kada je Zemljino magnetno polje mirno, usmjeravajući mokraću i izmet na osi sjever-jug.[42] Druga studija sugerira da psi mogu vidjeti Zemljino magnetno polje.[43][44]
Termoregulacija
[uredi | uredi izvor]
Psi prvenstveno reguliraju tjelesnu temperaturu dahtanjem [45] i znojenjem preko šapa. Dahtanjem pomjeraju hladni zrak preko vlažnih površina jezika i pluća, prenoseći toplotu u atmosferu.
Psi i drugi kanidi također imaju vrlo dobro razvijen skup nosnih turbinata, složeni skup kostiju i pripadajućih struktura mehkog tkiva (uključujući arterije i vene) u nosnoj šupljini. Te turbinate omogućavaju izmjenu toplote između malih arterija i vena na njihovim maksilloturbinatnim površinama (površine turbinata postavljene na maksila ma) u sistemu za izmjenu toplote protiv struje. Psi su sposobni za dugotrajne potjere, za razliku od grabežljivih mačaka u zasjedi, a ove složene turbine imaju važnu ulogu u omogućavanju toga (mačke imaju samo mnogo manji i manje razvijeni skup nosnih turbinata). Ova ista složena struktura zavoja pomaže u očuvanju vode u sušnim okruženjima. Sposobnosti očuvanja vode i termoregulacije u ovim dobro razvijenim zavojimaa kod pasa mogle su biti presudne prilagodbe koje su psima (uključujući i domaće pse i njihove divlje pretpovijesne pretke) omogućile preživljavanje u surovom okolišu Arktika i drugim hladnim područjima sjevera Evroazije i Sjeverne Amerike, koje su i vrlo suhe i vrlo hladne.
Reference
[uredi | uredi izvor]- ^ Scientists fetch useful information from dog genome publications Arhivirano 19. 11. 2020. na Wayback Machine, Cold Spring Harbor Laboratory, December 7, 2005; published online in Bio-Medicine
- ^ a b "World's Largest Dog". Arhivirano s originala, 19. 11. 2020. Pristupljeno 7. 1. 2008.
- ^ "Guinness World Records – Tallest Dog Living". Guinness World Records. 31. 8. 2004. Arhivirano s originala, 11. 7. 2011. Pristupljeno 7. 1. 2009.
- ^ Evans, Howard E.; de Lahunta, Alexander (2017). Guide to the Dissection of the Dog (8th izd.). St. Louis, Missouri: Elsevier. ISBN 9780323391658. OCLC 923139309.
- ^ Wayne, Robert K. (1986). "Cranial Morphology of Domestic and Wild Canids: The Influence of Development on Morphological Change". Evolution. 40 (2): 243–261. doi:10.2307/2408805. JSTOR 2408805. PMID 28556057.
- ^ Drake, Abby Grace; Klingenberg, Christian Peter (2010). "Large‐Scale Diversification of Skull Shape in Domestic Dogs: Disparity and Modularity". The American Naturalist. 175 (3): 289–301. doi:10.1086/650372. PMID 20095825.
- ^ Roberts, Taryn; McGreevy, Paul; Valenzuela, Michael (2010). "Human Induced Rotation and Reorganization of the Brain of Domestic Dogs". PLOS ONE. 5 (7): e11946. doi:10.1371/journal.pone.0011946. PMC 2909913. PMID 20668685. All cited in Roberts.
- ^ Roberts, Taryn; McGreevy, Paul; Valenzuela, Michael (2010). "Human Induced Rotation and Reorganization of the Brain of Domestic Dogs". PLOS ONE. 5 (7): e11946. doi:10.1371/journal.pone.0011946. PMC 2909913. PMID 20668685.
- ^ Christiansen, Per; Wroe, Stephen (2007). "Bite Forces and Evolutionary Adaptations to Feeding Ecology in Carnivores". Ecology. 88 (2): 347–358. doi:10.1890/0012-9658(2007)88[347:bfaeat]2.0.co;2. PMID 17479753.
- ^ Washington State University. "Respiratory System of the Dog". Pristupljeno 1. 6. 2017.[mrtav link]
- ^ Washington State University. "Digestive System of the Dog". Pristupljeno 31. 5. 2017.
- ^ a b Wang, Xiaoming and Tedford, Richard H. Dogs: Their Fossil Relatives and Evolutionary History. New York: Columbia University Press, 2008. pp.97-8
- ^ Sutter NB, Bustamante CD, Chase K, et al. (Apr 2007). "A single IGF1 allele is a major determinant of small size in dogs". Science. 316 (5821): 112–5. doi:10.1126/science.1137045. PMC 2789551. PMID 17412960.
- ^ Ostrander EA (Sep–Oct 2007). "Genetics and the Shape of Dogs; Studying the new sequence of the canine genome shows how tiny genetic changes can create enormous variation within a single species". Am. Sci. Arhivirano s originala, 26. 4. 2012. Pristupljeno 16. 11. 2020.
- ^ a b Klappenbach, Laura (2008). "What is Counter Shading?". About.com. Arhivirano s originala, 27. 9. 2011. Pristupljeno 22. 10. 2008.
- ^ a b Cunliffe, Juliette (2004). "Coat Types, Colours and Markings". The Encyclopedia of Dog Breeds. Paragon Publishing. str. 20–3. ISBN 0-7525-6561-3.
- ^ Candille SI, Kaelin CB, Cattanach BM, et al. (Nov 2007). "A -defensin mutation causes black coat color in domestic dogs". Science. 318 (5855): 1418–23. doi:10.1126/science.1147880. PMC 2906624. PMID 17947548.
- ^ Stanford University Medical Center, Greg Barsh et al. (2007, October 31). Genetics Of Coat Color In Dogs May Help Explain Human Stress And Weight. ScienceDaily. Retrieved September 29, 2008
- ^ "Genetics of Coat Color and Type in Dogs". Sheila M. Schmutz, Ph.D., Professor, University of Saskatchewan. 25. 10. 2008. Arhivirano s originala, 14. 2. 2020. Pristupljeno 5. 11. 2008.
- ^ "The Case for Tail Docking". cdb.org. Arhivirano s originala, 14. 4. 2009. Pristupljeno 22. 10. 2008.
- ^ "Dog Breeds Born Without Tails".
- ^ Ninomiya, Hiroyoshi; Akiyama, Emi; Simazaki, Kanae; Oguri, Atsuko; Jitsumoto, Momoko; Fukuyama, Takaaki (2011). "Functional anatomy of the footpad vasculature of dogs: Scanning electron microscopy of vascular corrosion casts". Veterinary Dermatology. 22 (6): 475–81. doi:10.1111/j.1365-3164.2011.00976.x. PMID 21438930.
- ^ a b c d e f Coren, Stanley (2004). How Dogs Think. First Free Press, Simon & Schuster. ISBN 0-7432-2232-6.
- ^ A&E Television Networks (1998). Big Dogs, Little Dogs: The companion volume to the A&E special presentation. A Lookout Book. GT Publishing. ISBN 1-57719-353-9.
- ^ a b Alderton, David (1984). The Dog. Chartwell Books. ISBN 0-89009-786-0.
- ^ a b c Jennifer Davis (1998). "Dr. P's Dog Training: Vision in Dogs & People". Arhivirano s originala, 9. 2. 2015. Pristupljeno 20. 2. 2015.
- ^ Anna A. Kasparson; Jason Badridze; Vadim V. Maximov (Jul 2013). "Colour cues proved to be more informative for dogs than brightness". Proceedings of the Royal Society B: Biological Sciences. 280 (1766): 20131356. doi:10.1098/rspb.2013.1356. PMC 3730601. PMID 23864600.
- ^ Jay Neitz; Timothy Geist; Gerald H. Jacobs (1989). "Color Vision in the Dog" (PDF). Visual Neuroscience. 3 (2): 119–125. doi:10.1017/s0952523800004430. PMID 2487095. Arhivirano s originala (PDF), 13. 4. 2015. Pristupljeno 23. 6. 2015.
- ^ Jay Neitz; Joseph Carroll; Maureen Neitz (Jan 2001). "Color Vision — Almost Reason Enough for Having Eyes" (PDF). Optics & Photonics News: 26–33. Arhivirano s originala (PDF), 4. 3. 2016. Pristupljeno 23. 6. 2015.
- ^ a b Mech, David. Wolves, Behavior, Ecology, and Conservation. The University of Chicago Press, 2006, p. 98.
- ^ Jonica Newby; Caroline Penry-Davey (25. 9. 2003). "Catalyst: Dogs' Eyes". Australian Broadcasting Corporation. Pristupljeno 26. 11. 2006.
- ^ Elert, Glenn; Timothy Condon (2003). "Frequency Range of Dog Hearing". The Physics Factbook. Pristupljeno 22. 10. 2008.
- ^ "How well do dogs and other animals hear". Pristupljeno 7. 1. 2008.
- ^ "How well do dogs and other animals hear".
- ^ "Dog Sense of Hearing". seefido.com. Arhivirano s originala, 1. 5. 2009. Pristupljeno 22. 10. 2008.
- ^ "Understanding a Dog's Sense of Smell". Dummies.com. Arhivirano s originala, 9. 3. 2008. Pristupljeno 22. 10. 2008.
- ^ "The Dog's Sense of" (PDF). Alabama and Auburn Universities. Arhivirano s originala (PDF), 11. 1. 2012. Pristupljeno 22. 10. 2008.
- ^ Virányi, Z. F.; Range, F. (2013). "Social learning from humans or conspecifics: Differences and similarities between wolves and dogs". Frontiers in Psychology. 4: 868. doi:10.3389/fpsyg.2013.00868. PMC 3849518. PMID 24363648.
- ^ Dijkgraaf S.; Vergelijkende dierfysiologie; Bohn, Scheltema en Holkema, 1978, ISBN 90-313-0322-4
- ^ Coren, Stanley
- ^ , Santos, A "Puppy and Dog Care: An Essential Puppy Training Guide", 2015 Amazon Digital Services, Inc. [1] Arhivirano 24. 6. 2015. na Wayback Machine
- ^ Hart, V.; Nováková, P.; Malkemper, E.; Begall, S.; Hanzal, V. R.; Ježek, M.; Kušta, T. Š.; Němcová, V.; Adámková, J.; Benediktová, K. I.; Červený, J.; Burda, H. (2013). "Dogs are sensitive to small variations of the Earth's magnetic field". Frontiers in Zoology. 10 (1): 80. doi:10.1186/1742-9994-10-80. PMC 3882779. PMID 24370002.
- ^ Nießner, Christine; Denzau, Susanne; Malkemper, Erich Pascal; Gross, Julia Christina; Burda, Hynek; Winklhofer, Michael; Peichl, Leo (2016). "Cryptochrome 1 in Retinal Cone Photoreceptors Suggests a Novel Functional Role in Mammals". Scientific Reports. 6: 21848. doi:10.1038/srep21848. PMC 4761878. PMID 26898837.
- ^ Magnetoreception molecule found in the eyes of dogs and primates Arhivirano 23. 12. 2016. na Wayback Machine MPI Brain Research, 22 February 2016
- ^ "Arhivirana kopija". Arhivirano s originala, 14. 12. 2013. Pristupljeno 16. 11. 2020.CS1 održavanje: arhivirana kopija u naslovu (link)
Dopunska literatura
[uredi | uredi izvor]- Klaus-Dieter Budras (7. 12. 2010). Anatomy of the Dog: With Aaron Horowitz and Rolf Berg. Schlütersche Verlagsgesellschaft mbH & Company KG. ISBN 978-3-89993-099-3.
- Horowitz, Alexandra (2009). Inside of a Dog: What Dogs See, Smell, and Know. New York: Charles Scribner's Sons. ISBN 9781416583400. OCLC 973655798. Inside of a Dog: What Dogs See, Smell, and Know.
- Howard E. Evans; Alexander de Lahunta (7. 8. 2013). Miller's Anatomy of the Dog - E-Book. Elsevier Health Sciences. ISBN 978-0-323-26623-9.
Vanjski linkovi
[uredi | uredi izvor]| Sisari |  | |
|---|---|---|
| Ostali kičmenjaci | ||
| Beskičmenjaci | ||
| Biljke | ||
| Discipline |
| |
| Ostale teme | ||
- ^ Miklósi, Adám (2009). Dog Behaviour, Evolution, and Cognition. Oxford University Press. doi:10.1093/acprof:oso/9780199295852.001.0001. ISBN 978-0-19-929585-2.
- ^ Xiaoming Wang (2008) Dogs: Their Fossil Relatives and Evolutionary History Columbia University Press. ISBN 9780231509435