Pampatheriidae
| Pampatheriidae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion mit Panzer von Holmesina | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mittleres Miozän bis Unteres Holozän | ||||||||||||
| 18 Mio. Jahre bis 10.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Pampatheriidae | ||||||||||||
| Paula Couto, 1954 | ||||||||||||
Die Pampatheriidae sind eine ausgestorbene Familie aus der Ordnung der Gepanzerten Nebengelenktiere, zu denen auch die heute noch bestehenden Gürteltiere gehören. Wie diese besaßen die Pampatherien einen Panzer, der den Oberteil des Kopfes, den Körper und den Schwanz bedeckte. Allerdings waren die Pampatherien deutlich größer als die Gürteltiere und erreichten ein Körpergewicht von bis zu 200 kg. Trotz ihrer äußerlichen Ähnlichkeit mit den Gürteltieren stellen die nächsten Verwandten aber die riesigen Glyptodontidae dar. Die Pampatherien entstanden höchstwahrscheinlich im Verlauf des Miozäns, die ältesten gesicherten Funde sind rund 18 Millionen Jahre alt. Ihr Ursprung liegt wie bei allen Nebengelenktieren in Südamerika. Im Verlauf des Pliozäns – mit der Entstehung einer Landbrücke nach Nordamerika – besiedelten sie auch nördlicher gelegene Regionen. Zum Ende des Pleistozäns starben die Pampatherien aus.
Im Vergleich zu den Glyptodonten und den Gürteltieren stellen die Pampatherien im Verlauf ihrer Stammesgeschichte eine eher formenarme Gruppe mit nur wenigen Vertretern dar. Sie bewohnten zumeist offenere sowie teils sehr trockene Landschaften und ernährten sich hauptsächlich pflanzlich. Der allgemeine Körperbau spricht eher gegen eine grabende Lebensweise, wie sie bei den heutigen Gürteltieren belegt ist. Allerdings ist es möglich, dass sie einzelne, von anderen Gräbern angelegte Baue weiternutzten. Insgesamt sind die Pampatherien aufgrund der eher seltenen Funde weitaus weniger gut erforscht als ihre Verwandten. Erstmals nachgewiesen wurden Vertreter der Pampatherien bereits in den 1830er Jahren mit Funden aus Höhlen im südlichen Brasilien. Der heute gültige Familienname stammt aus dem Jahr 1954.
Merkmale
[Bearbeiten | Quelltext bearbeiten]Allgemein und Körpergröße
[Bearbeiten | Quelltext bearbeiten]Die Vertreter der Pampatherien ähnelten äußerlich den heutigen Gürteltieren (Dasypoda). Sie besaßen einen Kopf mit leicht verlängerter Schnauze und einen Panzer, der sowohl den Oberkopf, den Körper als auch den Schwanz schützte. Jedoch waren sie deutlich größer als die Gürteltiere. So erreichten die größten Formen wie Holmesina und Pampatherium in der Spätphase ihrer Stammesgeschichte zum Ausgang des Pleistozän bis zu 3 m Gesamtlänge[1][2] und wogen verschiedenen Berechnungen zufolge zwischen 176 und 195 kg bei ersteren sowie 214 und 240 kg bei letzteren Vertretern.[3][4] Ein vollständiges Skelett von Holmesina aus der Gruta da Lapinha im östlichen Brasilien weist eine Gesamtlänge von 2,2 m auf.[5] Frühere Formen wie Vassallia waren mit einer Körperlänge von rund 1 m und einem Gewicht von 60 bis 90 kg kleiner[6] und besaßen etwa die Größe des Riesengürteltiers, das der größte Angehörige der heutigen Gürteltiere ist. Dieses besitzt im Vergleich dazu eine Gesamtlänge von 1,5 m und ein Gewicht von rund 50 kg.[7] Aufgrund ihrer Größe und ihrer Panzergestaltung werden die Pampatherien teilweise auch giant armadillos („Riesengürteltiere“) genannt.[8][9]
Schädel- und Gebissmerkmale
[Bearbeiten | Quelltext bearbeiten]
Der Schädel der Pampatherien gleicht in etwa dem der Gürteltiere, vor allem dem der euphractinen Formen mit näherer Beziehung zum Sechsbinden-Gürteltier (Euphractus sexinctus), war aber massiver, höher und breiter gebaut. Er wurde bei großen Vertretern zwischen 28 und 36 cm lang.[10] Die Stirnlinie verlief häufig nach oben aufgewölbt, bei Kraglievichia bestand aber eine leichte Eindellung. Das Rostrum war lang ausgezogen, was deutlich von den nahe verwandten Glyptodontidae mit ihren stark verkürzten Schädeln abweicht. Dadurch erreichte das Nasenbein mit 14 bis 16 cm auch eine enorme Länge. Hinter den Augenfenstern befand sich eine starke Einschnürung des Schädels. Die Jochbögen luden seitlich weit aus. Sie besaßen einen kräftigen Bau mit hohen Knochen. In der Seitenansicht waren der vordere und hintere Bogenansatz stark nach unten orientiert, beide trafen sich etwa auf der Höhe der oberen Zahnreihe. Hier war die Oberfläche stark aufgeraut und markiert so Ansatzstellen für kräftige Muskeln. Auf den Scheitelbeinen bestand ein deutlich ausgeprägter Scheitelkamm.[11][12]
Auch der Unterkiefer besaß einen massigen Bau und war deutlich kräftiger als bei den Gürteltieren. Eine Verknöcherung der Symphyse kam nur bei sehr alten Individuen vor, ansonsten waren die beiden Unterkieferhälften in der Regel nicht fest verwachsen. Der Unterkieferkörper zeigte am Unterrand einen deutlich nach unten geschwungenen Verlauf, was an die Glyptodonten erinnert. Dadurch war der Unterkiefer hier auch sehr hoch. Die Gelenkenden reichten sehr weit nach oben. Das Gebiss wich wie bei allen Nebengelenktieren in seinem strukturellen Aufbau von dem der meisten Höheren Säugetiere ab. Je Kieferhälfte bestanden sowohl im Ober- als auch im Unterkiefer 9 Zähne, somit setzte sich das Gebiss aus insgesamt 36 Zähnen zusammen. Alle ausgebildeten Zähne werden als molarenartig (molariform) betrachtet, Schneide- und Eckzähne sowie Prämolaren waren nicht vorhanden. Im Unterschied zu der jeweils homodonten Zahngestaltung der Gürteltiere und der Glyptodonten war das Gebiss bei den Pampatherien heterodont aufgebaut. Die vorderen Zähne bildeten eine kurze Reihe und waren nagelartig gestaltet mit rundlich-ovalem Querschnitt. Die hinteren wiesen eine deutliche Längsstreckung auf und besaßen an den jeweiligen Enden zwei quergestellte, lappen- oder zungenartige Verbreiterungen (Loben), was den Zähnen einen bilobaten Charakter gab. Den Zähnen fehlte der harte Zahnschmelz, was als allgemein üblich für die Nebengelenktiere gilt. Stattdessen bestanden sie aus Zahnbein, wobei im Inneren überwiegend harter Orthodentin ausgebildet war. Bei einigen Vertretern verlief entlang der Mittelachse der hinteren Zähne eine Rippe aus besonders hartem Osteodentin auf der Kaufläche, die im Bereich der Loben verdickt war. Die Verteilung der harten Bestandteile des Zahnbeins variierte jedoch innerhalb der einzelnen Vertreter der Pampatherien stark, was als Anpassung an bestimmte Ernährungsweisen gedeutet wird. Bei einigen Vertretern waren die hinteren Zähne in der Seitenansicht stufenartig angeordnet.[11][12][13][14] Die Länge der Zahnreihe im Oberkiefer erreichte bis zu 20 cm.[10]
Körperskelett
[Bearbeiten | Quelltext bearbeiten]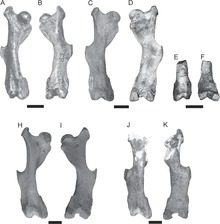
Die Wirbelsäule eines vollständigen Individuums setzte sich aus 7 Hals-, 10 Brust-, 1 Lenden-, 11 Kreuzbein- und 22 Schwanzwirbeln zusammen.[5] Besonderheiten des Körperskeletts finden sich vor allem an der Wirbelsäule. Die Halswirbel waren wie bei den Gürteltieren und den Glyptodonten auch miteinander verwachsen und bildeten eine knöcherne Röhre. Dies betrifft weitgehend den Axis (zweiter Halswirbel) und die darauf folgenden dritten bis fünften Halswirbel, während der Atlas (erster Halswirbel) frei beweglich stand. Eine besondere Ausnahme stellt dabei Vassallia dar, ein Vertreter der Pampatherien aus dem Oberen Miozän, bei dem der Axis ebenfalls frei beweglich und nicht mit der Halsröhre verwachsen war.[15] Typisch für alle Nebengelenktiere sind zusätzliche Gelenkflächen (xenarthrische Gelenke oder Xenarthrale) an den seitlichen Fortsätzen der hinteren Brustwirbel und an den Lendenwirbeln.[16] Der Bewegungsapparat zeigte nur wenige Besonderheiten. Am Mittelschaft des Oberschenkelknochens trat ein sehr prominenter Dritter Rollhügel (Trochanter) auf, der als Muskelansatzstelle fungierte und typisch für Nebengelenktiere ist; bei den verwandten Glyptodontidae saß dieser deutlich weiter unten nahe dem Knie. Schien- und Wadenbein waren miteinander verwachsen. Hände und Füße zeigten sich weitgehend unspezialisiert, der innere und äußere Finger der Hand war jeweils in seiner Länge reduziert, die drei mittleren Finger besaßen jedoch keine deutlichen Längsstreckungen wie bei den Gürteltieren. Alle Strahlen trugen Krallen.[17][18]
Panzer
[Bearbeiten | Quelltext bearbeiten]

Wie alle Gepanzerten Nebengelenktiere besaßen auch die Pampatherien eine äußere, knöcherne Panzerung bestehend aus kleinen Knochenplättchen (Osteoderme). Der Panzer bedeckte den Oberteil des Schädels, den Rumpf und den Schwanz. Er setzte sich bei großen Vertretern wie Holmesina aus über 850 Einzelelementen zusammen, wovon mehr als 630 auf den Körperpanzer, über 50 auf den Kopf und rund 130 auf den Schwanz entfallen.[19] Der Körperpanzer ähnelte dem der heutigen Gürteltiere und bestand aus einem festen Schulter- und Beckenteil mit dazwischen liegenden, beweglichen Bändern. Abweichend von den Gürteltieren mit ihrer variablen Anzahl an beweglichen Bändern wiesen die Pampatherien immer drei auf. Die starren und beweglichen Teile des Panzers waren aus quer verlaufenden Reihen von kleinen Knochenplättchen aufgebaut, die wohl eine Hornbedeckung trugen.[20] Die Knochenplättchen des starren Panzers besaßen eine längliche, meist recht- oder mehreckige Form und wiesen auf der Oberfläche im Gegensatz zur teils komplexen Musterung der Gürteltiere und Glyptodonten nur eine zentrale, flache Erhebung auf, die von einer hufeisenförmig gestalteten Rille umgeben war, bei Vassallia und Pampatherium erschien sie nahezu flach.[9] An den beweglichen Bändern waren die Knochenplättchen langgestreckt geformt und setzten sich aus zwei Abschnitten zusammen, einem kurzen Basisteil und einer längeren „Zunge“. Auf der Oberfläche befanden sich meist mehrere tiefe Löcher. Aufgrund der einfachen Oberflächengestaltung wird für die Pampatherien angenommen, dass nur eine Hornplatte ein Osteoderm überdeckte und nicht mehrere wie bei den Gürteltieren, deren Knochenplättchen eine komplexe Musterung auf der Oberfläche tragen.[15][21] Der Schwanz war ebenfalls vollständig von einem Panzer umgeben. Die Knochenplättchen hier waren analog zum Rückenpanzer in Reihen angeordnet, die sich aber im Gegensatz zu den Gürteltieren nicht überlappten, sondern dicht aneinanderstießen.[22][19]
Im Querschnitt wiesen die Osteoderme einen dreilagigen Aufbau auf. Oben und unten befand sich je eine feste Knochenschicht, die jeweils einen Bereich umschlossen, welcher aus zahlreichen Hohlräumen bestand. Er nahm etwa die Hälfte der Dicke des Knochenplättchens ein. In diese spongiose Struktur waren Schweißdrüsen und Haarfollikel eingelagert. Die feste Knochenschicht bestand abwechselnd aus harten Lagen und kollagenreichen Faserbündeln (Sharpeysche Fasern). Eine Besonderheit der Knochenplättchen der Pampatherien zeigte sich in dem Vorkommen und der Ausprägung dieser Faserbündel auch an den Seitenrändern der Plättchen des steifen Panzers, wo diese mit den benachbarten Plättchen verwachsen waren. Bei den Gürteltieren haben die Sharpeyschen Fasern in diesem Bereich eine andere Ausrichtung, bei Glyptodonten wurden sie bisher nicht beobachtet. Insgesamt gab der Aufbau der Knochenplättchen diesen eine gewisse Elastizität und schützte sie vor starken, von außen einwirkenden Kräften.[23][24]
Verbreitung und Lebensraum
[Bearbeiten | Quelltext bearbeiten]Pampatherien waren ursprünglich auf Südamerika beschränkt, wo sie sich wenigstens seit dem Mittleren Miozän vor rund 14 Millionen Jahren entwickelten. Sie sind hier hauptsächlich in den Tiefländern der Pamparegion im zentralen Bereich des Kontinents bis zum Amazonasbecken nachgewiesen, im letzteren war ihre Verbreitung aber eher gering. Außerdem kamen sie in den westlichen Küstenregionen vor. Im Gegensatz zu den verwandten Glyptodonten und den heutigen Gürteltieren gibt es aber bisher keine Überlieferung aus den Hochlagen der Anden.[25] Ab dem Mittleren Pliozän besiedelten die Pampatherien durch die Entstehung der Landbrücke nach Nordamerika auch nördlichere Regionen. Fossilfunde liegen hauptsächlich aus den südlichen und südöstlichen Bereichen der USA von Texas über die Golfküste bis nach Florida vor. Sehr weit nördlich gelegene Fundstellen sind in Kansas zu finden.[26] Die Pampatherien waren an offenere Landschaften angepasst und besiedelten sowohl die wüstenartig kargen Steppengebiete der Pampa als auch feuchtere Küstenregionen. Häufig treten Pampatherien und Glyptodonten gemeinsam im Fossilbericht auf. Dabei sind beide nicht als ökologisch identisch, sondern nur als ähnlich einzustufen.[27][9]
Paläobiologie
[Bearbeiten | Quelltext bearbeiten]Fortbewegung
[Bearbeiten | Quelltext bearbeiten]Im Bezug auf den Bewegungsapparat stehen die Pampatherien vermittelnd zwischen den kleineren Gürteltieren und den teils riesigen Glyptodonten. Die Hinterbeine sind wie bei den Glyptodonten an einen schwerfälligen Gang angepasst. An den Vorderbeinen sind keine stärkeren Spezialisierungen auffällig. So ist der obere Gelenkfortsatz der Elle, das Olecranon, vergleichsweise kürzer als bei den Gürteltieren mit ihren teils außerordentlich langen Fortsätzen. Ein massives Olecranon ist häufig ein Indikator für eine grabende Lebensweise, wie es bei den meisten Gürteltieren der Fall ist, da dieses die Hebelwirkung des Trizeps verstärkt, der als Strecker des Unterarms fungiert. Auch die Ausprägung der Hand spricht eher gegen deutliche grabende Tätigkeiten, da die drei Mittelfinger (II bis IV) nicht verlängert sind und keine kräftigen, zum Bodenwühlen geeigneten Klauen ausgebildet waren. Gürteltiere haben als aktive Gräber häufig stark verlängerte Mittelfinger mit stark gebogenen und ebenfalls verlängerten Grabkrallen. Die anatomischen Merkmale der Pampatherien lassen also keine deutliche Anpassung an eine unterirdische (fossoriale) Lebensweise erkennen.[18][28] Über die allgemeine Gliedmaßenstruktur hinaus ermöglicht der Bau des Innenohrs Rückschlüsse auf die Beweglichkeit eines Tieres und den Gleichgewichtssinn. Hier sind hauptsächlich die Bogengänge von Bedeutung, deren Weite Auswirkungen auf die Agilität hat. Das bisher untersuchte Innenohr von Holmesina weist eher enge Bogengänge auf, die allerdings weiter und dünner sind als im Vergleich zu den Glyptodonten, sie entsprechen denen eines größeren Teils der heutigen Gürteltiere. Daher wird eine ähnliche Agilität wie bei letzteren angenommen, wobei als direkter Vergleich das Riesengürteltier in Frage kommt. Der jeweilige Winkel zwischen den drei Bogengängen ist kleiner als 90°. Dies stimmt mit zahlreichen grabenden Tieren einschließlich der Gürteltiere überein und impliziert eine geringere Rotationsfähigkeit des Kopfes.[29]
Allerdings sind aus der Miramar-Formation in der Gegend um Mar del Plata an der Ostküste Argentiniens Grabbaue bekannt, die in das Untere und Mittlere Pleistozän datieren. Während größere Strukturen zwischen 0,9 und 1,8 m Weite und teilweise bis zu 40 m Länge anhand der Größe und der charakteristischen, zweilinigen Kratzspuren auf große Bodenfaultiere aus der Gruppe der Mylodontidae zurückgeführt werden können, sind einzelne kleinere von rund 1 m Weite und 75 cm Höhe mit anderen Kratzspuren versehen. Diese weisen generell drei Linien von maximal 30 cm Länge auf. Sie entsprechen in etwa denen der Vorderfüße der Pampatherien und könnten daher auf diese zurückgeführt werden. Auch die Größe der Baue entspricht in etwa der der Pampatherien, deren Panzer einen Durchmesser von 80 cm erreichen konnte. Sie sind auch deutlich größer als die des heutigen Riesengürteltiers.[30][28] Bei anderen, ähnlich großen Strukturen aus dem südlichen Brasilien wurden zusätzlich Abdrücke der Osteoderme an den Wänden gefunden, die aber eher auf große Gürteltiere wie Propraopus hinweisen. Es ist jedoch nicht ausgeschlossen, dass derartige Baue nach dem Verlassen des Erbauers von anderen Tieren wie eben auch Pampatherien weiter benutzt wurden.[31]
Ernährungsweise
[Bearbeiten | Quelltext bearbeiten]Die Zähne der Pampatherien sind teilweise relativ hochkronig (hypsodont), was im Vergleich mit anderen Säugetiergruppen eher auf eine Bevorzugung harter, abrasiver Grasnahrung hinweist. Da den Zähnen der Zahnschmelz fehlt, sind Untersuchungen zur genauen Ernährungsweise, etwa in Form von Isotopenanalysen, jedoch nicht möglich. Heutige Gürteltiere, die mit stiftförmigen Zähnen ausgestattet sind, umfassen sowohl Alles- als auch Fleischfresser, unter ausgestorbenen Formen sind auch einzelne Pflanzenfresser bekannt. Spezielle Untersuchungen des Kauapparates zeigen, dass die Pampatherien Ähnlichkeiten zu den heutigen Huftieren mit pflanzenfresserischer Ernährungsweise besitzen. Dazu gehört unter anderem der hohe Gelenkansatz am Unterkiefer, der die Hebelwirkung beim Kauen verstärkte und so eine höhere Beißkraft erlaubte. Dies steht auch im Verhältnis zu den deutlich weit ausladenden Jochbögen mit ihrer stark aufgerauten unteren Knochenausstülpung, was eine wesentlich stärkere Kaumuskulatur, vor allem des hier ansetzenden Musculus masseter annehmen lässt. Weiterhin ist die Gelenkfläche des Unterkiefers eher flach gestaltet und unterstützte so horizontale Kaubewegungen. Darauf weisen auch häufig quer verlaufende Abrasionsspuren auf den mittleren Zähnen hin. Wie bei den heutigen Pflanzenfressern sind die mittleren Zähne zudem stark verlängert und die gesamte Zahnreihe deutlich nach hinten versetzt. An den hinteren molarenartigen Zähnen ist zusätzlich im Zentrum der Kaufläche härteres Zahnbein ausgebildet, was die Schereffizienz beim Kauen erhöht. Alle diese Merkmale, die sich schon bei den früheren Vertretern wie Kraglievichia und Vassallia nachweisen lassen, deuten darauf hin, dass die Pampatherien an pflanzliche Nahrung – möglicherweise Gräser – angepasst waren. Die Hauptunterschiede innerhalb der Pampatherien finden sich dabei in der Ausprägung der Muskulatur, wobei dieses mit der paläobiogeographischen Verbreitung der einzelnen Formen und dem daraus resultierenden unterschiedlichen Nahrungsangebot in Verbindung gebracht wird. So sind einige Arten, die in der eher trockenen Pamparegion vorkamen, an weit härtere und widerstandsfähigere Pflanzen angepasst gewesen, während solche aus der feuchteren Küstenregion des heutigen Ecuadors und Perus, die damals wohl von Savannen oder Trockenwäldern geprägt waren, weniger abrasive Pflanzennahrung bevorzugten. Allgemein werden die Pampatherien dadurch als auf primär grashaltige Nahrung spezialisiert angesehen mit sekundärer, landschaftsabhängiger Anpassung an festere oder faserigere Pflanzenteile.[12][11][32] Bemerkenswert ist zudem, dass die Pampatherien im Bezug auf ihre Körpergröße eine relativ große Kaufläche aller Zähne zusammengenommen aufweisen, die im Vergleich zu den nahe verwandten Glyptodontidae wesentlich umfangreicher ist. So besaß Holmesina bei einem Körpergewicht von 177 kg eine Kaufläche von etwa 1520 mm², was knapp unterhalb der der heutigen Tapire liegt, die bei 210 bis 290 kg Körpergewicht zwischen 1530 und 2240 mm² zur Verfügung haben. Im Vergleich dazu wies der Glyptodonten-Vertreter Plohophorus, der rund 260 kg wog, mit 900 mm² nur fast die Hälfte an Kaufläche auf. Möglicherweise führte die weniger komplexe Zahngestaltung der Pampatherien gegenüber den Glyptodonten und die Reduktion an scharfkantigen Rippen auf den Zahnoberflächen zu einer Vergrößerung der Kaufläche und damit einer besseren Verwertung der aufgenommenen Nahrung im Mundraum.[33]
Gehirn
[Bearbeiten | Quelltext bearbeiten]Von Pampatherium konnte ein vollständiger Ausguss der Gehirnkapsel untersucht werden. Diese umfasste ein Volumen von rund 133 cm³ bei einem vermuteten Körpergewicht des Tieres von 210 kg. Der Enzephalisationsquotient lag somit bei 0,27. Für das ältere Vassallia wird ein Wert von 0,17 angenommen. Beides liegt in der Variationsbreite der Glyptodontidae (0,12 bis 0,4), aber im unteren Skalenbereich der Gürteltiere (0,44 bis 1,06). Vergleichbar den Glyptodonten und Gürteltieren war der Riechkolben ausgesprochen groß und nahm etwa 8 % des Gesamtvolumens des Gehirns ein. Zwei Drittel beanspruchte des Weiteren das Großhirn, der Rest verfiel auf das Kleinhirn. Der Riechkolben hatte eine schmal-spitze Form, was dem der Glyptodonten entsprach. Dagegen besitzen die Gürteltiere einen eher breiten Riechkolben. Am relativ einfach gestalteten Relief der Großhirnrinde war lediglich der Sulcus suprasylvianus des Scheitellappens erkennbar, der bei den Gürteltieren zusätzlich ausgebildete Sulcus praesylvianus fehlte hingegen.[34][35]
Systematik
[Bearbeiten | Quelltext bearbeiten]Äußere Systematik
[Bearbeiten | Quelltext bearbeiten]
Verwandtschaftsverhältnis der Pampatherien und Gürteltiere laut morphologischen Daten nach Billet et al. 2011[36]
|

Die Pampatheriidae stellen eine Familie innerhalb der Ordnung der Gepanzerten Nebengelenktiere (Cingulata) dar. Dieser einst formenreich in Süd- und Nordamerika verbreiteten Ordnung gehören weiterhin auch die ebenfalls ausgestorbenen Glyptodontidae, Peltephilidae, die Pachyarmatheriidae und Palaeopeltidae sowie die heute noch bestehenden Gürteltiere (Dasypoda) an. Die Gepanzerten Nebengelenktiere werden wiederum zur Überordnung der Nebengelenktiere (Xenarthra) gezählt, welche zusätzlich noch die Zahnarmen (Pilosa) mit den heutigen Faultieren (Folivora) und den Ameisenbären (Vermilingua) einschließt. Die Nebengelenktiere repräsentieren eine der vier Hauptlinien der Höheren Säugetiere, die den anderen drei (zusammengefasst als Epitheria) als Schwestergruppe gegenübersteht.[37] Eine generelle Gemeinsamkeit der Nebengelenktiere besteht in den namengebenden xenarthrischen Gelenken (Nebengelenke, auch Xenarthrale) an den Gelenkfortsätzen der hinteren Brust- und der Lendenwirbel. Eine Ausnahme bilden hier allerdings die Glyptodontidae, deren Lendenwirbel als einzigartiges Merkmal miteinander verwachsen sind.[38] Der Ursprung der Nebengelenktiere ist bisher nicht geklärt, die ältesten Fossilfunde, die zu den Gürteltieren gerechnet werden, stammen aus Südamerika und gehören dem Paläozän vor mehr als 56 Millionen Jahren an.[39] Aufgrund molekulargenetische Untersuchungen ist eine Abspaltung der Nebengelenktiere von den anderen Höheren Säugetieren bereits in der ausgehenden Unterkreide vor etwa 103 Millionen Jahren wahrscheinlich. Die Gürteltiere als heute lebende, nächste Verwandte der Pampatherien trennten sich von der gemeinsamen Linie mit den Zahnarmen zu Beginn des Paläozäns vor etwa 65 Millionen Jahren ab.[40][41]
Die nächsten Verwandten der Pampatherien innerhalb der Gepanzerten Nebengelenktiere sind die Glyptodonten. Diese zeichnen sich durch einen vollständig starren Rückenpanzer aus und erreichten zudem mit bis zu 2 t Körpergewicht eine enorme Größe.[42] Aufgrund des charakteristischen Baus des Panzers mit den drei beweglichen Bändern hielt man die Pampatherien ursprünglich für näher verwandt mit den Gürteltieren; sie wurden innerhalb dieser in einer eigenen Unterfamilie geführt. Allerdings zeigten Untersuchungen an Schädeln und Zähnen eine nähere Beziehung der Pampatherien zu den Glyptodonten. Unter anderem konnte dies an der Struktur des Gehörganges und am Bau des Kauapparates, wie dem hohen Unterkiefer, belegt werden. Darüber hinaus sind bei den Vertretern beider Familien die Zähne komplexer gestaltet. Bei den Gürteltieren treten einfache, nagelartig gebaute Zähne auf, bei den Pampatherien und den Glyptodonten sind diese deutlich verlängert und durch zwei beziehungsweise drei querstehende Loben charakterisiert.[2][43] Die Pampatherien und Glyptodonten bilden zusammen die übergeordnete Gruppe der Glyptodonta. Als Schwestergruppe der Glyptodonta erwiesen sich die Pachyarmatheriidae, deren Kennzeichen ein sich überlappender Rückenpanzer ohne bewegliche Bänder dazwischen ist. Sie waren vom Oberen Miozän bis zum beginnenden Holozän in Süd- und Nordamerika verbreitet.[44] Die Glyptodonta selbst sind skelettanatomischen Untersuchungen aus den Jahren 2006 und 2011 zufolge tiefer in die Gruppe der Gürteltiere eingebettet, da sie möglicherweise eine engere Bindung an die Euphractinae aufweisen, also der Gruppe um das Sechsbinden-Gürteltier, das Zwerggürteltier und die Borstengürteltiere. Gemeinsam mit diesen formen sie eine evolutionäre Entwicklungslinie.[2][36] Im Kontrast dazu zeigten im Jahr 2016 veröffentlichte molekulargenetische Studien an Doedicurus aus der Gruppe der Glyptodonten eine nähere Beziehung zu den Tolypeutinae (Kugelgürteltiere, Nacktschwanzgürteltiere und Riesengürteltier) und den Chlamyphorinae (Gürtelmulle). Inwiefern die Pampatherien hier eine ähnliche Position einnehmen wie die Glyptodonten ist unklar, da bisher noch kein Erbgut gewonnen wurde. Allerdings unterstützt die Morphologie des Innenohrs einerseits die enge Beziehung der Pampatherien zu den Glyptodonten, andererseits auch die genetisch gewonnene Erkenntnis der Nahverwandtschaft letzterer mit den Chlamyphorinae.[29] In beiden Untersuchungsergebnissen, also sowohl den skelettanatomischen als auch den molekulargenetischen, bilden die Gürteltiere eine paraphyletische Gruppe.[45][46] Die weiteren Gruppen wie die Palaeopeltidae und Peltephilidae stehen in einem mehr oder weniger engen Verhältnis zu den übrigen Gepanzerten Nebengelenktieren.[36][47]
Innere Systematik
[Bearbeiten | Quelltext bearbeiten]Die Pampatherien gelten als eher konservative Gruppe, die kaum Entwicklungen durchlief und dadurch insgesamt weniger variantenreich erscheint als ihre nahen Verwandten, die Gürteltiere und Glyptodonten. Unterfamilien werden innerhalb der Familie der Pampatherien nicht unterschieden. Generell erfolgt die Identifizierung der Gattungen und Arten ähnlich wie bei den anderen Gruppen der Gepanzerten Nebengelenktiere anhand der unterschiedlichen Ausbildung der Osteoderme des Panzers und nicht wie sonst üblich bei Säugetieren mit Hilfe von Schädel- und Skelettmerkmalen. Grund dafür ist die relativ große Häufigkeit der Knochenplättchen im Vergleich mit Skelettmaterial.[48] Anhand der Knochenplättchen lassen sich innerhalb der Gruppe zwei Entwicklungslinien feststellen. Die eine umfasst Vassallia und Pampatherium, die beide durch wenig ornamentierte Panzerplättchen ausgezeichnet sind, die andere kann mit Plaina, Kraglievichia und Holmesina angenommen werden.[9]
Überblick über die Gattungen
[Bearbeiten | Quelltext bearbeiten]Es sind etwa ein halbes Dutzend Gattungen anerkannt:[49][50][51]
- Familie: Pampatheriidae (=Chlamydotheriidae) Paula Couto, 1954
- Scirrotherium Edmund & Theodor, 1997
- Kraglievichia Castellanos, 1927
- Vassallia Castellanos, 1927
- Plaina Castellanos, 1937
- Tonnicinctus Góis, González Ruiz, Scillato-Yané & Soibelzon , 2015
- Pampatherium (=Chlamytherium, Chlamydotherium, Hoffstetteria) Gervais & Ameghino, 1880
- Holmesina Simpson, 1930
Als unsicher zu den Pampatherien gehörig wird folgende Gattung angesehen:[50]
- Machlydotherium Ameghino, 1902
Die Abgrenzung der einzelnen Gattungen wird unterschiedlich bewertet. Einige Forscher sehen Plaina auch als synonym zu Kraglievichia.[49][15] In der Vergangenheit wurde Holmesina teilweise als identisch zu Pampatherium aufgefasst.[52] Generell wird eine Revision der Familie angemahnt.[48]
Stammesgeschichte
[Bearbeiten | Quelltext bearbeiten]Ursprünge
[Bearbeiten | Quelltext bearbeiten]Der Ursprung der Pampatherien liegt wie der der anderen Nebengelenktiere höchstwahrscheinlich in Südamerika. Als ältester Vertreter gilt teilweise Machyldotherium aus dem Oberen Eozän vor 35 bis 37 Millionen Jahren. Funde stammen unter anderen aus Gran Barranca südlich des Sees Colhué Huapi im nördlichen Patagonien.[53][54] Weitere Fossilreste in Form von Knochenplättchen entstammen der etwas jüngeren Sarmiento-Formation im nordöstlich vom Colhué-Huapi-See gelegenen Barrancas Blancas, ebenfalls Patagonien,[55] sowie aus der Guabirotuba-Formation im südbrasilianischen Bundesstaat Paraná.[56] Die Stellung von Machyldotherium innerhalb der Pampatherien ist aber umstritten.[57][9]
Miozän
[Bearbeiten | Quelltext bearbeiten]
Durch die unsichere Zuweisung von Machyldotherium gilt das erst 1997 beschriebene Scirrotherium als bisher unzweifelhaft ältestes Mitglied der Linie der Pampatherien. Zu den frühesten Nachweisen gehört ein Knochenplättchen aus der Castillo-Formation im nordwestlichen Venezuela, deren Alter auf 19 bis 17 Millionen Jahre und damit in das Untere Miozän datiert.[58][59] Umfangreicheres Material stammt aus der La-Venta-Formation in Kolumbien und besteht aus einem nur 14 cm langen Schädel mit Unterkiefer und zusätzlich noch zahlreichen Resten des Körperskelettes sowie einigen Knochenplättchen. Ihr Alter liegt bei etwa 14 bis 15 Millionen Jahren, was dem Mittleren Miozän entspricht.[60] Die Funde verweisen auf einen sehr kleinen Angehörigen mit nur etwa 40 kg Körpergewicht.[61][9] Erst für den Beginn des Oberen Miozäns ist die Gattung auch für die südlicher gelegenen Bereiche Südamerikas belegt. Hierzu gehören Funde aus der Paraná-Formation in der argentinischen Provinz Entre Ríos und der Puerto-Madryn-Formation im äußersten Nordosten der argentinischen Provinz Chubut. Beide Gesteinseinheiten sind eigentlich durch marine Sedimente geprägt, enthalten unter anderem mit Knochenplättchen von Scirrotherium auch einzelne terrestrische Elemente. Die Funde werden auf ein Alter von rund 8,7 Millionen Jahren geschätzt.[62][63] Herausragend ist der sehr fossilreiche Conglomerodo ossifero der Ituzaingó-Formation, die durch den Río Paraná in Entre Ríos aufgeschlossen wird. Die umfangreichen Reste der Nebengelenktiere enthalten unter anderem auch zahlreiche Osteoderme von Scirrotherium, das hier seinen forschungsgeschichtlich frühesten Nachweis außerhalb des nördlichen Südamerikas hat.[64][50] Daneben konnten hier aber auch Schädelreste und Teile des Körperskeletts von Kraglievichia aufgedeckt werden, einem weiteren frühen und relativ kleinen Vertreter.[65] Ebenfalls zu diesen frühen Formen gehört Vassallia, das anhand eines Unterkiefers und einiger Knochenplättchen aus der Andalhuala-Formation in der argentinischen Provinz Catamarca beschrieben wurde. Dieser Gattung kann auch ein nahezu vollständiger Schädel aus der Corral-Quemado-Formation in der gleichen Provinz zugeschrieben werden.[15]
Noch vor dem Einsetzen des Großen Amerikanischen Faunenaustausches, der durch die Entstehung der Landbrücke zwischen Süd- und Nordamerika mit der Schließung des Isthmus von Panama im Verlauf des Pliozäns ausgelöst wurde, sind Pampatherien im Oberen Miozän nördlich des südamerikanischen Kontinents zu finden. Der Curré-Formation im südlichen Costa Rica entstammen Knochenplättchen und Reste des Körperskeletts von Scirrotherium. Möglicherweise bestanden vor der Kollision der beiden Kontinentalmassen kleinere Inseln oder Inselbögen, die von den Pampatherien besiedelt wurden.[66]
Pliozän und Pleistozän
[Bearbeiten | Quelltext bearbeiten]

Das Pliozän ist vor allem durch die weitere Ausbreitung der Pampatherien nach Norden geprägt. Aus Ablagerungen im Becken von San Miguel Allende im zentralmexikanischen Bundesstaat Guanajuato sind Osteoderme überliefert, die auf 4,7 bis 4,8 Millionen Jahre datiert werden und der Gattung Plaina angehören. Im nachfolgenden Pleistozän sind dann nur noch die terminalen Formen Holmesina, Pampatherium und Tonnicinctus überliefert, letzteres ist aber bisher auf nur zwei Fundstellen beschränkt.[67][68][51]
Holmesina tritt erstmals vor etwa 2,0 bis 2,6 Millionen Jahren am Übergang vom Pliozän zum Pleistozän in Nordamerika in Erscheinung. Es wird angenommen, dass die Gattung dort auch aus vorhergehenden Einwanderern wie Plaina entstanden ist. Zu den ältesten Funden gehört ein nahezu vollständiges Skelett von der bedeutenden Fossilfundstelle Haile, 6 km nordöstlich von Newberry im Alachua County in Florida. Die Funde kamen genauer in der Lokalität 15A, einer sedimentgefüllten Kalksteinspalte, zum Vorschein.[69] Dabei waren die ersten Immigranten in Nordamerika noch relativ klein, sie wogen rund 44 kg bei einer Oberschenkelknochenlänge von etwa 20 cm. Bis zum Ende des Pleistozäns nahmen sie um ein Vielfaches an Körpergewicht zu und wogen dann etwa 220 kg. Diese große Formen besaßen dann Oberschenkelknochen mit einer Länge von 33 cm.[4] Holmesina kehrte während des Pleistozäns auch nach Südamerika zurück, wo es hauptsächlich im Oberen Pleistozän belegt ist. Hier durchlief es offenbar eine erneute Differenzierung und ist daher mit mehreren Arten überliefert. Unter anderem aus der Gruta da Lapinha bei Iramaia im brasilianischen Bundesstaat Bahia sind zwei vollständige Skelette geborgen worden.[9][5] Wann genau diese Rückwanderung begann, ist unbekannt; in El Salvador sind Reste von Holmesina in Form von Knochenplättchen bereits aus dem Übergang vom Alt- zum Mittelpleistozän bekannt.[70]
Für Pampatherium wird dagegen angenommen, dass es sich im Unteren Pleistozän in Südamerika aus Formen wie Vassallia entwickelte. Es vollzog ähnlich wie Holmesina eine massive Größenentwicklung zum ausgehenden Pleistozän hin. Möglicherweise erreichte es in einer weiteren Auswanderungswelle auch Mittel- und Nordamerika. Allerdings ist es nicht so weit nach Norden vorgedrungen wie sein Verwandter; die nördlichsten Funde stammen aus dem Oberpleistozän von Mexiko, etwa aus Térapa. Gründe für die stärker begrenzte Ausbreitung von Pampatherium könnten in der besseren Anpassung an deutlich trockenere Klimaverhältnisse der Gattung liegen.[8]
Aussterben
[Bearbeiten | Quelltext bearbeiten]Sowohl Pampatherium als auch Holmesina starben zum Ende des Pleistozäns im Zuge der quartären Aussterbewelle aus. Junge Daten für Pampatherium stammen aus dem brasilianischen Bundesstaat Bahia und liegen bei rund 13.000 BP.[9] Mit 13.600 bis 14.400 BP nur wenig älter sind Radiocarbondaten für Holmesina aus La Brea im Nordwesten Perus. Aus der weiter südlich davon gelegenen Wüste Cupisnique an der peruanischen Westküste, von wo relativ umfangreiches Skelettmaterial von Holmesina stammt, liegen Altersdaten vor, die zwischen 8.900 und 25.000 BP schwanken.[25] Die quartäre Aussterbewelle in Amerika fällt zusammen mit der Erstbesiedlung beider Kontinente durch frühe Jäger und Sammler, was vor etwa 15.000 Jahren begann. Ob aktive Bejagung zum Verschwinden der Pampatherien führte, ist unklar. In Nordamerika sind von rund zwei Dutzend Fundstellen aus der Endphase des Pleistozäns, die Reste von Pampatherien enthalten, nur wenige bekannt, die in Zusammenhang mit frühmenschlicher Jagd stehen.[71][72] Dazu gehört Lubbock Lake in Texas, eine bedeutende, 120 ha große Fundstelle in einem Altarm des Yellowhouse Draw, einem Nebenfluss des Brazos River. Die unterste Fundschicht wird auf 11.100 Jahre datiert und enthielt neben zahlreichen Knochenresten auch Steingeräte, vor allem Schlagsteine und steinerne Arbeitsunterlagen (Ambosse). Die Knochenreste, darunter auch von Holmesina, zeigen Schlagmarken und geben so eine Verwendung als Nahrungsgrundlage an. Die Funde von Holmesina am Lubbock Lake gehören zu den jüngsten bekannten der Pampatherien überhaupt.[73][74]
Forschungsgeschichte
[Bearbeiten | Quelltext bearbeiten]
Die Erforschung der Pampatherien reicht bis in die erste Hälfte des 19. Jahrhunderts zurück. Sie gehören damit zu den ersten, ehemals endemisch in Amerika verbreiteten Tieren, die der Wissenschaft bekannt wurden. Im Jahr 1836 hatte der dänische Paläontologe Peter Wilhelm Lund (1801–1880) Reste von Pampatherien in Höhlen bei Lagoa Santa im brasilianischen Bundesstaat Minas Gerais entdeckt und bereits im November 1837 zwei Vorberichte veröffentlicht, wobei er auch bereits erste Funde von Nebengelenktieren erwähnte.[75] Eine umfangreichere Abhandlung war 1839 zu Papier gebracht, die jedoch erst weitere zwei Jahre später erscheinen sollte. In einem zwischenzeitlich von Hans Christian Ørsted zusammengestellten Überblicksartikel zu Lunds Forschungen erwähnte dieser die neue Gattung Chlamytherium.[76] Lunds eigener Bericht wurde dann 1841 veröffentlicht. In diesem stellte er die Gattung Chlamydotherium vor, das Material, das ihm für seine Beschreibung zur Verfügung stand, umfasste einige Knochenplättchen und Kieferfragmente.[77][22] (Bereits 1839 erschienen in mehreren bedeutenden, wissenschaftlichen Zeitschriften Europas, etwa in den Ausgaben des Londoner Natural History Museum[78] oder des Pariser Muséum national d’histoire naturelle[79] Kurzberichte zu Lunds Arbeiten, die dieser brieflich übermittelt hatte und in denen er Chlamydotherium als tapirgroßes Gürteltier ebenfalls erwähnt.) Der Gattungsname Chlamydotherium setzt sich aus den griechischen Wörtern χλαμύς (chlamys „Mantel“, Genitiv χλαμύδος chlamydos) und θηρίον (thēríon „Tier“) zusammen. Jedoch hatte Heinrich Georg Bronn bereits im Jahr 1838 einen Vertreter der Glyptodontidae, nahen Verwandten der Pampatherien, anhand von Funden aus Brasilien und Uruguay mit dem Namen Chlamydotherium versehen,[80] wodurch dieser präokkupiert war (er gilt aber heute als synonym zu Glyptodon).[10][81][52]
In der zweiten Hälfte des 19. Jahrhunderts war über ausgestorbene Gürteltiere und Pampatherien im Gegensatz zu den Glyptodonten kaum etwas bekannt. Im Jahr 1875 erwähnt der argentinische Forscher Florentino Ameghino (1854–1911) in einem Manuskript erstmals Pampatherium, wohl in Unkenntnis von Lunds Chlamytherium beziehungsweise Chlamydotherium. Seine Publikation ist aber nicht mehr erhalten, möglicherweise auch nie erschienen. Allerdings erkannte er zusammen mit Henri Gervais im Jahr 1880, dass Pampatherium identisch ist mit Lunds Chlamydotherium (dass der Name durch einen Glyptodontenvertreter präokkupiert ist, erkennen beide Autoren zu diesem Zeitpunkt nicht). Aufgrund des Aufbaus des Panzers mit beweglichen Bändern heben sie die enge Verbindung von Chlamydotherium zu den Gürteltieren hervor, bemerken aber durch den Aufbau der Zähne des Unterkiefers auch Gemeinsamkeiten mit den Glyptodonten.[82] Chlamytherium findet keine Erwähnung in der Schrift, in der Folgezeit bis weit in das 20. Jahrhundert hinein wurde dieses häufig als Gattungsname weiter verwendet.[10][52]
Die ersten Funde in Form von Knochenplättchen in Nordamerika wurden von Joseph Willcox im Jahr 1888 am Peace Creek in Florida entdeckt. Er leitete diese an den amerikanischen Paläontologen Joseph Leidy (1823–1891) weiter, der sie im Jahr darauf als zu Glyptodon gehörig beschrieb. Noch im gleichen Jahr, nachdem er die Übereinstimmungen in der Gestaltung der Knochenplättchen erkannte, korrigierte Leidy die Gattungszugehörigkeit zu Lunds Chlamytherium. Weitere Fossilreste folgten ebenfalls aus Florida und aus Texas und umfassten neben den Osteodermen auch Unterkiefer und Schädelteile.[83] Diese zeigten auf, dass teilweise Unterschiede zu den südamerikanischen Formen bestehen. Im Jahr 1930 benannte deswegen George Gaylord Simpson (1902–1984) unter Berufung auf einen Unterkiefer und einzelner Oberkieferfragmente aus Bradenton Field in Florida die Gattung Holmesina.[84]
Bereits 1889 hatte Ameghino den Familiennamen Chlamydotheriidae geprägt, worin dessen Erkenntnis zu Grunde liegt, das Pampatherium nur ein Synonym zu Lunds Chlamydotherium sei. Im Jahr 1930 änderte Oliver Perry Hay diesen in Chlamytheriinae ab, wobei er neben der Namenskorrektur die Gruppe auf die Ebene der Unterfamilie innerhalb der Gürteltiere verschob.[85] Carlos de Paula Couto (1910–1982) wies dann 1954 darauf hin, dass der von Ameghino 1875 benutzte Name Pampatherium valide ist und gegenüber Chlamytherium vorzuziehen sei, da letzterer Name in dem Moment ungültig wurde, als Lund beschloss, ihn in Chlamydotherium umzubenennen. Sein daraufhin vorgeschlagener Name Pampatheriidae für die Familie setzte sich in der Folgezeit langsam durch.[69]
Einzelnachweise
[Bearbeiten | Quelltext bearbeiten]- ↑ Gordon Edmund: The fossil giant armadillos of North America (Pampatheriinae, Xenarthra Edentata). In: G. Gene Montgomery (Hrsg.): The Ecology and Evolution of Armadillos, Sloths, and Vermilinguas. Smithsonian Institution Press, 1985, S. 83–93
- ↑ a b c Timothy J. Gaudin und John R. Wible: The Phylogeny of Living and Extinct Armadillos (Mammalia, Xenarthra, Cingulata): A Craniodental Analysis. In: M. T. Carrano, T. J. Gaudin, R. W. Blob und J. R. Wible (Hrsg.): Amniote Paleobiology. Chicago/London: University of Chicago Press, 2006, S. 153–198
- ↑ Fernando H. De S. Barbosa, Laís Alves-Silva, Alexandre Liparini und Kleberson De Oliveira Porpino: Reviewing the body size of some extinct Brazilian Quaternary Xenarthrans. Journal of Quaternary Science 39 (8), 2024, S. 1160–1167, doi:10.1002/jqs.3560
- ↑ a b H. Gregory McDonald: Paleoecology of extinct Xenarthrans and the Great American Biotic Interchange. Bulletin of the Florida Museum of Natural History 45 (4), 2005, S. 313–333
- ↑ a b c Jorge Felipe Moura, Flávio Góis, Fernando Carlos Galliari und Marcelo Adorna Fernandes: A new and most complete pampathere (Mammalia, Xenarthra, Cingulata) from the Quaternary of Bahia, Brazil. Zootaxa 4661 (3), 2019, S. 401–444, doi:10.11646/zootaxa.4661.3.1
- ↑ Darin A. Croft und Velizar Simeonovski: Horned armadillos and rafting monkeys. The fascinating fossil mammals of South America. Indiana University Press, 2016, S. 1–304 (S. 196–197)
- ↑ Arnaud Leonard Jean Desbiez und Danilo Kluyber: The Role of Giant Armadillos (Priodontes maximus) as Physical Ecosystem Engineers. Biotropica 45 (5), 2013, S. 537–540
- ↑ a b Jim I. Mead, Sandra L. Swift, Richard S. White, H. Greg McDonald und Arturo Baez: Late Pleistocene (Rancholabrean) Glyptodont and Pampathere (Xenarthra, Cingulata) from Sonora, Mexico. Revista Mexicana de Ciencias Geológica 24 (3), 2007, S. 439–449
- ↑ a b c d e f g h G. J. Scillato-Yané, A. A. Carlini, E. P. Tonni und J. I. Noriega: Paleobiogeography of the late Pleistocene pampatheres of South America. Journal of South American Earth Sciences 20, 2005, S. 131–138
- ↑ a b c d Flávio Góis, Gustavo Juan Scillato-Yané, Alfredo Armando Carlini und Martín Ubilla: Una nueva especie de Holmesina Simpson (Xenarthra, Cingulata, Pampatheriidae) del Pleistoceno de Rondônia, Sudoeste de la Amazonia, Brasil. Revista Brasileira de Paleontologia 15 (2), 2012, S. 211–227
- ↑ a b c Gerardo De luliis, María S. Bargo und Sergio F. Vizcaíno: Variation in skull morphology and mastication in the fossil giant armadillos Pampatherium spp. and allied genera (Mammalia: Xenarthra: Pampatheriidae), with comments on their systematics and distribution. Journal of Vertebrate Paleontology 20 (4), 2000, S. 743–754
- ↑ a b c Sergio F. Vizcaíno, Gerardo De luliis und María S. Bargo: Skull shape, masticatory apparatus, and diet of Vassalliaand Holmesina (Mammalia: Xenarthra: Pampatheriidae). When anatomy constrains destiny. Journal of Mammalian Evolution 5 (4), 1998, S. 291–322
- ↑ Sergio F. Vizcaíno: The teeth of the “toothless”: novelties and key innovations in the evolution of xenarthrans (Mammalia, Xenarthra). Paleobiology 35 (3), 2009, S. 343–366
- ↑ Daniela C. Kalthoff: Microstructure of Dental Hard Tissues in Fossil and Recent Xenarthrans (Mammalia: Folivora and Cingulata). Journal of Morphology 272, 2011, S. 641–661
- ↑ a b c d Gerardo De Iuliis und A .Gordon Edmund: Vassallia maxima Castellanos, 1946 (Mammalia: Xenarthra: Pampatheriidae), from Puerta del Corral Quemado (Late Miocene to Early Pliocene), Catamarca Province, Argentina. Robert J. Emry (Hrsg.): Cenozoic mammals of land and sea: Tributes to the Career of Clayton E. Ray. Smithonia Contributions to Paleobiology 93, 2002, S. 49–63
- ↑ Timothy J. Gaudin: The Morphology of Xenarthrous Vertebrae (Mammalia: Xenarthra). Fieldiana Geology, New Series 41, 1999, S. 1–38
- ↑ H. Gregory McDonald: Xenarthran skeletal anatomy: primitive or derived? Senckenbergiana biologica 83, 2003, S. 5–17
- ↑ a b N. Milne, Sergio. F. Vizcaíno und J. C. Fernicola: A 3D geometric morphometric analysis of digging ability in the extant and fossil cingulate humerus. Journal of Zoology 278 (1), 2009, S. 48–56
- ↑ a b Gordon Edmund: The Armor of the fossil giant armadillos (Pampatheriidae, Xenarthra, Mammalia). Texas Memorial Museum, The University of Texas, Austin, 1985, S. 1–20
- ↑ Santiago Rodriguez-Bualó, Alfredo Eduardo Zurita, Flávio Góis, Ángel R. Miño-Boilini, Esteban Soibelzon und Freddy Paredes-Ríos: Pampatheriidae (Xenarthra, Cingulata) from the Tarija valley, Bolivia: A taxonomic update. Rivista Italiana di Palrontologia e Stratigrafia 120 (2), 2014, S. 253–259
- ↑ Thais Matos Pereira Ferreira, Daniel de Melo Casali, Simone Baes Das Neves und Ana Maria Ribeiro: Osteoderm morphology and taxonomy of Pampatheriidae (Cingulata, Xenarthra) from the Quaternary of the Neotropical region. Historical Biology, 2024, doi:10.1080/08912963.2024.2439939
- ↑ a b Gideon T. James: An Edentate from the Pleistocene of Texas. Journal of Paleontology 31 (4), 1957, S. 796–808
- ↑ Robert V. Hill: Comparative Anatomy and Histology of Xenarthran Osteoderms. Journal of Morphology 267, 2005, S. 1441–1460
- ↑ Dominik Wolf, Daniela C. Kalthoff und P. Martin Sander: Osteoderm Histology of the Pampatheriidae (Cingulata, Xenarthra, Mammalia): Implications for Systematics, Osteoderm Growth, and Biomechanical Adaptation. Journal of Morphology 273, 20912, S. 388–404
- ↑ a b François Pujos und Rodolfo Salas: A systematic reassessment and paleogeographic review of fossil Xenarthra from Peru. Bulletin de l'Institut Français d'Etudes Andines 33 (2), 2004, S. 331–377
- ↑ Claude W. Hibbard, Richard J. Zakrzewski, Ralph E. Eshelman, Gordon Edmund, Clayton D. Griggs und Caroline Griggs: Mammals from the Kanopolis Local Fauna, Pleistocene (Yarmouth) of Ellsworth County, Kansas. Museum of Paleontology, University of Michigan 25 (2), 1978, S. 11–48
- ↑ David D. Gillette und Clayton E. Ray: Glyptodonts of North America. Smithonian Contributions to Paleobiology 40, 1981, S. 1–251
- ↑ a b Sergio F. Vizcaíno, Marcelo Zaráte, M. Susana Bargo und Alejandro Dondas: Pleistocene burrows in the Mar del Plata area (Argentina) and their probable builders. Acta Palaeontologica Polonica 46 (2), 2001, S. 289–301
- ↑ a b P. Sebastián Tambusso, Luciano Varela, Flávio Góis, Jorge Felipe Moura, Chiara Villa und Richard A. Fariña: The inner ear anatomy of glyptodonts and pampatheres (Xenarthra, Cingulata): Functional and phylogenetic implications. Journal of South American Earth Sciences 108, 2021, S. 103189, doi:10.1016/j.jsames.2021.103189
- ↑ Alejandro Dondas, Federico I. Isla und José L. Carballido: Paleocaves exhumed from the Miramar Formation (Ensenadan Stage-age, Pleistocene), Mar del Plata, Argentina. Quaternary International 210, 2009, S. 44–50
- ↑ Francisco Sekiguchi Buchmann, Renato Pereira Lopes und Felipe Caron: Paleotoca do Município de Cristal, RS. Registro da atividade fossorial de mamíferos gigantes extintos no sul do Brasil. In: M. Winge, C. Schobbenhaus, C. R. G. Souza, A. C. S. Fernandes, M. Berbert-Born, W. Sallun filho und E. T. Queiroz: (Hrsg.): Sítios Geológicos e Paleontológicos do Brasil. 2010, S. 1–10
- ↑ Sergio F. Vizcaíno, Richard A. Fariña, M. Susana Bargo und Gerardo De Iuliis: Functional and phylogenetic assessment of the masticatory adaptations in Cingulata (Mammalia, Xenarthra). Ameghiniana 41 (4), 2004, S. 651–664
- ↑ Sergio F. Vizcaíno, M. Susana Bargo und Guillermo H. Cassini: Dental occlusal surface area in relation to body mass, food habits and other biological features in fossil xenarthrans. Ameghiniana 43 (1), 2006, S. 11–26
- ↑ P. Sebastián Tambusso und Richard A. Fariña: Digital cranial endocast of Pseudoplohophorus absolutus (Xenarthra, Cingulata) and its systematic and evolutionary implications. Journal of Vertebrate Paleontology 35 (5), 2015, S. e967853, doi:10.1080/02724634.2015.967853
- ↑ P. Sebastián Tambusso und Richard A. Fariña: Digital endocranial cast of Pampatherium humboldtii (Xenarthra, Cingulata) from the Late Pleistocene of Uruguay. Swiss Journal of Palaeontology 134, 2015, S. 109–116, doi:10.1007/s13358-015-0070-5
- ↑ a b c Guillaume Billet, Lionel Hautier, Christian de Muizon and Xavier Valentin: Oldest cingulate skulls provide congruence between morphological and molecular scenarios of armadillo evolution. Proceedings of the Royal Society B, 278, 2011, S. 2791–2797
- ↑ Maureen A. O’Leary, Jonathan I. Bloch, John J. Flynn, Timothy J. Gaudin, Andres Giallombardo, Norberto P. Giannini, Suzann L. Goldberg, Brian P. Kraatz, Zhe-Xi Luo, Jin Meng, Xijun Ni, Michael J. Novacek, Fernando A. Perini, Zachary S. Randall, Guillermo W. Rougier, Eric J. Sargis, Mary T. Silcox, Nancy B. Simmons, Michelle Spaulding, Paúl M. Velazco, Marcelo Weksler, John R. Wible und Andrea L. Cirranello: The Placental Mammal Ancestor and the Post–K-Pg Radiation of Placentals. Science 339, 2013, S. 662–667, doi:10.1126/science.1229237
- ↑ Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, S. 1–431 (S. 200–204)
- ↑ Sergio F. Vizcaíno und W. J. Loughry: Xenarthran biology: Past, present and future. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 1–7.
- ↑ Frédéric Delsuc, Michael J. Stanhope und Emmanuel J.P. Douzery: Molecular systematics of armadillos (Xenarthra, Dasypodidae): contribution of maximum likelihood and Bayesian analyses of mitochondrial and nuclear genes. Molecular Phylogenetics and Evolution 28, 2003, S. 261–275
- ↑ Frédéric Delsuc, Sergio F Vizcaíno und Emmanuel J. P. Douzery: Influence of Tertiary paleoenvironmental changes on the diversification of South American mammals: a relaxed molecular clock study within xenarthrans. BMC Evolutionary Biology 4 (11), 2004, S. 1–13
- ↑ Leopold Héctor Soibelzon, Martín Zamorano und Gustavo Juan Scillato-Yané: Un Glyptodontidae de gran tamaño en el Holoceno Temprano de la Region Pampeana, Argentinia. Revista Brasileira de Paleontologia 15 (1), 2012, S. 105–112
- ↑ Juan-Carlos Fernicola, Serdio F. Vizcaíno und Richard a. Fariña: The evolution of armored xenarthrans and the phylogeny of the glyptodonts. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 79–85
- ↑ Juan C. Fernicola, Andrés Rinderknecht, Washington Jones, Sergio F. Vizcaíno und Kleberson Porpino: A new species of Neoglyptatelus (Mammalia, Xenarthra, Cingulata) from the late Miocene of Uruguay provides new insights on the evolution of the dorsal armor in cingulates. Ameghiniana 55, 2018, S. 233–252
- ↑ Frédéric Delsuc, Gillian C. Gibb, Melanie Kuch, Guillaume Billet, Lionel Hautier, John Southon, Jean-Marie Rouillard, Juan Carlos Fernicola, Sergio F. Vizcaíno, Ross D. E. MacPhee und Hendrik N. Poinar: The phylogenetic affinities of the extinct glyptodonts. Current Biology 26, 2016, S. R155–R156, doi:10.1016/j.cub.2016.01.039
- ↑ Kieren J. Mitchell, Agustin Scanferla, Esteban Soibelzon, Ricardo Bonini, Javier Ochoa und Alan Cooper: Ancient DNA from the extinct South American giant glyptodont Doedicurus sp. (Xenarthra: Glyptodontidae) reveals that glyptodonts evolved from Eocene armadillos. Molecular Ecology 25 (14), 2016, S. 3499–3508, doi:10.1111/mec.13695
- ↑ Daniel Perera, Pablo Toriño und Martín Ciancio: La Presencia del Xenartro Palaeopeltis inornatus Ameghino, 1894, en la Formación Fray Bentos (Oligoceno Tardío), Uruguay. Ameghiniana 51 (3), 2014, S. 254–258
- ↑ a b Richard M. Fariña, Sergio F. Vizcaíno und Gerardo de Iuliis: Megafauna. Giant beasts of Pleistocene South America. Indiana University Press, 2013, S. 1–436 (S. 220–223), ISBN 978-0-253-00230-3
- ↑ a b Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 86–87)
- ↑ a b c Flávio Góis, Gustavo Juan Scillato-Yané, Alfredo Armando Carlini und Edson Guilherme: A new species of Scirrotherium Edmund & Theodor, 1997 (Xenarthra, Cingulata, Pampatheriidae) from the late Miocene of South America. Alcheringa: An Australasian Journal of Palaeontology 37 (2), 2013, S. 177–188
- ↑ a b Flávio Góis, Laureano Raúl González Ruiz, Gustavo Juan Scillato-Yané und Esteban Soibelzon: A Peculiar New Pampatheriidae (Mammalia: Xenarthra: Cingulata) from the Pleistocene of Argentina and Comments on Pampatheriidae Diversity. PlosOne 10 (6), 2015, S. e0128296 doi:10.1371/journal.pone.0128296
- ↑ a b c Castor Cartelle Guerra und Germán Arturio Bohórquez Mahecha: Pampatherium paulacoutoi, uma nova espécie de tatu gigante da Bahia, Brasil (Edentata, Dasypodidae). Revista Brasileira de Zoologia, 2, 1984, S. 229–254
- ↑ Richard F. Kay, Richard H. Madden, M. Guiomar Vucetich, Alfredo A. Carlini, Mario M. Mazzoni, Guillermo H. Re, Matthew Heizleri und Hamish Sandeman: Revised geochronology of the Casamayoran South American Land Mammal Age: Climatic and biotic implications. PNAS 96 (23), 1999, S. 13235–13240
- ↑ Alfredo A. Carlini, M. Ciancio und Gustavo J. Scillato-Yané: Los Xenarthra de Gran Barranca, más de 20 Ma de historia. Congreso Geológico Argentino 16 (4), 2005, S. 419–424
- ↑ María Teresa Dozo, Martín Ciancio, Pablo Bouza und Gastón Martínez: Nueva asociación de mamíferos del Paleógeno en el este de la Patagonia (provincia de Chubut, Argentina): implicancias biocronológicas y paleobiogeográficas. Andean Geology 41 (1), 2014, S. 224–247, doi:10.5027/andgeoV41n1-a09.
- ↑ Fernando A. Sedor, Édison V. Oliveira, David D. Silva, Luiz A. Fernandes, Renata F. Cunha, Ana M. Ribeiro und Eliseu V. Dias: A New South American Paleogene Land Mammal Fauna, Guabirotuba Formation (Southern Brazil). Journal of Mammalian Evolution 24, 2017, S. 39–55
- ↑ Robert Hoffstetter: Remarques sur la phylogénie et la classification des édentés xénarthres (mammifères) actuels et fossiles. Bulletin du Muséum National d'Histoire Naturelle, 2nd série 41 (1), 1969, S. 91–103
- ↑ Ascanio D. Rincón, Andrés Solórzano und H. Gregory McDonald: New mammal records from the Early Miocene Castillo Formation of Venezuela. Journal of Vertebrate Paleontology 28 (3; suppl), 2010, S. 151a
- ↑ Ascanio D. Rincón, Andrés Solórzano, Mouloud Benammi, Patrick Vignaud und H. Gregory McDonald: Chronology and geology of an Early Miocene mammalian assemblage in North of South America, from Cerro La Cruz (Castillo Formation), Lara State, Venezuela: implications in the ‘changing course of Orinoco River’ hypothesis. Andean Geology 41 (3), 2014, S. 507–528, doi:10.5027/andgeoV41n3-a02.
- ↑ A. Gordon Edmund und Jessica Theodor: A new giant pampatheriid armadillo. In: Richard F. Kay, Richard H. Madden, Richard L. Cifelli und John J. Flynn (Hrsg.): Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, 1997, S. 227–232
- ↑ Alfredo A. Carlini, Sergio F. Vizcaíno und Gustavo J. Scillato-Yané: Armored Xenarthtans: A unique taxonomic and ecologic assemblage. In: Richard F. Kay, Richard H. Madden, Richard L. Cifelli und John J. Flynn (Hrsg.): Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, 1997, S. 213–226
- ↑ Adriana M. Candela, Ricardo A. Bonini und Jorge I. Noriega: First continental vertebrates from the marine Parana´ Formation (Late Miocene, Mesopotamia, Argentina): Chronology, biogeography, and paleoenvironments. Geobios 45, 2012, S. 515–526
- ↑ María Teresa Dozo, Pablo Bouza, Alejandro Monti, Luis Palazzesi, Viviana Barreda, Gabriela Massaferro, Roberto A. Scasso und Claudia P. Tambussi: Late Miocene continental biota in Northeastern Patagonia (Península Valdés, Chubut, Argentina). Palaeogeography, Palaeoclimatology, Palaeoecology 297, 2010, S. 100–109
- ↑ Alberto Luis Cione, María de las Mercedes Azpelicueta, Mariano Bond, Alfredo A. Carlini, Jorge R. Casciotta, Mario Alberto Cozzuol, Marcelo de la Fuente, Zulma Gasparini, Francisco J. Goin, Jorge Noriega, Gustavo J. Scillato-Yané, Leopoldo Soibelzon, Eduardo Pedro Tonni, Diego Verzi und María Guiomar Vucetich: Miocene vertebrates from Entre Ríos province, eastern Argentina. In: F. G. Aceñolaza und R. Herbst (Hrsg.): El Neógeno de Argentina. Universidad Nacional de Tucumán, 2000, S. 191–237
- ↑ Gustavo J. Scillato-Yané, Flávio Góis, Alfredo E. Zurita, Alfredo A. Carlini Laureano R. Gonzáles Ruiz, Cecilia M. Krmpotic, Cristian Oliva und Martín Zamorano: Los Cingulata (Mammalia, Xenarthra) del “Conglomerado osífero” (Mioceno Tardío) de la Formación Ituzaingó de Entre Ríos, Argentina. In: Diego Brandoni und Jorge I. Noriega (Hrsg.): El Neógeno de la Mesopotamia argentina. Buenos Aires, 2013, S. 118–134
- ↑ César A. Laurito und Ana L. Valerio: Scirrotherium Antelucanus, una nueva especie de Pampatheriidae (Mammalia, Xenarthra, Cingulata) del Mioceno Superior de Costa Rica, América Central.- Revista Geológica de América Central 49, 2013, S. 45–62, doi:10.15517/rgac.v0i49.13101
- ↑ Oscar Carranza Castañeda und Wade E. Miller: Late Tertiary terrestrial mammals from Central Mexico and their relationship to South American immigrants. Revista Brasileira de Paleontologia 7 (2), 2004, S. 249–261
- ↑ Michael O. Woodburne: The Great American Biotic Interchange: Dispersals, Tectonics, Climate, Sea Level and Holding Pens. Journal of Mammal Evolution 17, 2010, S. 245–264
- ↑ a b Jesse Steadman Robertson: Latest Pliocene mammals from Haile 15A, Alachua County, Florida. Bulletin of the Florida State Museum, Biological Sciences 20, 1976, S. 111–186 (S. 124–142)
- ↑ Daniel H. Aguilar und César A. Laurito: El armadillo gigante (Mammalia, Xenarthra, Pampatheriidae) del Río Tomayate, Blancano tardío-Irvingtoniano temprano, El Salvador, América Central. Revista Geológica de América Central 41, 2009, S. 25–36, doi:10.15517/rgac.v0i41.4176
- ↑ Donald K. Grayson und David J. Meltzer: Clovis Hunting and Large Mammal Extinction: A Critical Review of the Evidence. Journal of World Prehistory 16 (4), 2002, S. 313–359
- ↑ J. Tyler Faith und Todd A. Surovell: Synchronous extinction of North America’s Pleistocene mammals. PNAS 106, 2009, S. 20641–20645
- ↑ Eileen Johnson und Vance T. Holliday: Lubbock Lake: Late Quaternary cultural and environmental change on the Southern High Plains, USA. Journal of Quaternary Science 4 (2), 1989, S. 145–165
- ↑ Vance T. Holliday: Late Pleistocene and Holocene stratigraphy, Southern High Plains of Texas. Geological Society of America, Centennial Field Guide - South-Central Section 1988, S. 293–298
- ↑ Peter Wilhelm Lund: Om Huler i Kalksteen i det indre af Brasilien, der tildeels indeholde fossile knokler. Anden Afhandling. Lapa de Cerca Grande. Det Kongelige Danske Videnskabernes Selskbas Naturvidenskabelige og Matematiske Afhandlinger 6, 1837, S. 207–249 und 309–332 ([1])
- ↑ Hans Christian Ørsted: Oversigt over det Kongelike Danske Videnskabernes Selskabs Forhandlinger og dets Medlemmers Arbeiter i Aaret 1838. Kopenhagen, 1839, S. 1–32 (S. 11) ([2])
- ↑ Peter Wilhelm Lund: Blik paa Brasiliens Dyreverden för Sidste Jordomvaeltning. Anden Afhandling: Patteedyrene. Det Kongelige Danske Videnskabernes Selskbas Naturvidenskabelige og Matematiske Afhandlinger 8, 1839, S. 61–144 (S. 69) ([3])
- ↑ Peter Wilhelm Lund: List of fossil Mammifera from the Basin of Rio das Velhas, with an extract of some of their distinguishing Characters. Annals of Natural History 3, 1839, S. 422–427 ([4])
- ↑ Peter Wilhelm Lund: Coup d’oeil sur les espèces éteintes de mammifères du Brésil; extrait de quelques mémoires presentés à l'Académie Royale des Sciences de Copenhague. Annales des sciences naturelles, Zoologie 2 (11), 1939, S. 214–234 ([5])
- ↑ Heinrich Georg Bronn: Lethaea geognostica: oder, Abbildungen und Beschreibung der für die Gebirgs-Formationen bezeichnendsten Versteinerungen, Band 2. Stuttgart, 1838, S. 1–1346 (S. 1256–1259) ([6])
- ↑ Cástor Cartelle: Peter W. Lund, a naturalist of several sciences. Lundiana 3 (2), 2002, S. 83–85
- ↑ Henri Gervais und Florentino Ameghino: Les mammifères fossiles de l´Ámérique du Sud. Paris-Buenos Aires, 1880, S. 1–225 (S. 210–211)
- ↑ Alvin R. Cahn: Chlamytherium septentrionalis, a Fossil Edentate New to the Fauna of Texas. Journal of Mammalogy 3 (1), 1922, S. 22–24
- ↑ George Gaylord Simpson: Holmesina septentrionalis, extinct giant armadillo of Florida. American Museum Novitates 442, 1930, S. 1–10
- ↑ Oliver Perry Hay: Second bibliography and catalogue of the fossil Vertebrata of North America. Volume II. Carnegie Institution, Washington, 1930, S. 1–1074 (S. 417–418)