Gunneraceae
| Gunneráceas | ||
|---|---|---|
.jpg) Gunnera manicata | ||
| Taxonomía | ||
| Reino: | Plantae | |
| División: | Magnoliophyta | |
| Clase: | Eudicotyledoneae | |
| Orden: | Gunnerales | |
| Familia: |
Gunneraceae Meisn., 1842, nom. cons. | |
| Sinonimia | ||
ver texto
| ||
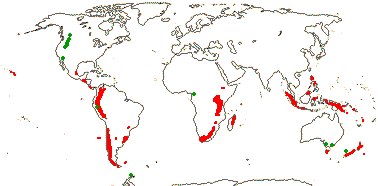
Las gunneráceas (Gunneraceae) son una familia de Angiospermas del Orden Gunnerales. Tan solo consta del género Gunnera L. con 62 especies, presentando una distribución relíctica disyunta, principalmente en el hemisferio sur, con escasos representantes en el hemisferio norte.
Algunas de sus especies ostentan el récord de mayor tamaño de hoja en una dicotiledónea, llegando a alcanzar e incluso sobrepasar los 4 m de ancho, por lo que son muy apreciadas en jardinería.

Son plantas monoicas, bisexuales o polígamas, a veces hermafroditas, raramente dioicas. Son hierbas perennes, a veces gigantescas (hasta de 6 m de altura), en un caso (Gunnera herteri) anual, con o sin estolones, con glándulas mucilaginosas, con órganos especializados (raíces adventicias abortadas o ficorrizas) verdeazulados u oscuros de aspecto glandular conteniendo células endosimbióticas de cianobacterias del género Nostoc.
Sus hojas son simples, pecioladas, alternas, envainantes, sin estípulas, limbos oblongos a reniformes o bien peltados, los márgenes dentados, crenados o palmatilobados, la base cuneada o cordada, con hidatodos prominentes, nervadura en las hojas grandes palinactinódroma y con los nervios costiformes en el envés y actinódroma o pinnada en las hojas pequeñas, con los nervios poco prominentes; hipostomáticas o anfistomáticas. Estomas anomocíticos. Posee tallos gruesos, con las hojas en el ápice y cicatrices foliares en la parte inferior, en las especies de gran tamaño, o cortos y erectos con una roseta de hojas, formando céspedes, en las especies de tamaño pequeño, a veces con numerosas escamas más o menos visibles, simples o divididas entre las bases foliares, estolones con un par de brácteas o solo una en forma de ócrea en el ápice, sistema vascular polistélico en los tallos y sifonostélico en los estolones, con un número de haces vasculares relacionado con el tamaño de los tallos (3-5 en las especies de tallos finos, 50-60 en las de tallos medianos, y varios cientos en las de tallo grueso), nudos multilacunares con numerosos rastros foliares (unilacunares en Gunnera herteri). Se ha encontrado, sin embargo, un sistema vascular eustélico en los tallos y protostélico en el rizoma de Gunnera insignis.[1]
Sus inflorescencias son axilares o pseudoterminales, a veces escapiformes, erectas, usualmente en racimos simples o compuestos, en espigas o en panículas, las flores superiores masculinas, las inferiores femeninas, las medianas a veces perfectas, o todas unisexuales. Las flores son pequeñas, sin bráctea pero con bracteolas, epíginas, perfectas, disimétricas (con un eje anteroposterior y otro transversal) y dímeras (raramente trímeras). Sépalos 2, libres, valvados, en el eje anteroposterior, a veces escasamente visibles. Pétalos 2, libres, cuculados, caducos, apenas más largos que los sépalos, en el eje transversal, ausentes de las flores femeninas. Androceo epipétalo o libre, de (1-)2 estambres iguales, fértiles, con filamentos cortos, situados en el eje transversal, anteras ditecas, tetrasporangiadas, dehiscencia latrorsa por hendiduras longitudinales. Gineceo ínfero, de 2 carpelos, sincárpico, unilocular, estilos 2, libres, apicales, subulados o comprimidos, papilosos a plumosos, en el eje transversal, estigmas papilosos, secos, óvulo solitario, péndulo, anátropo, bitégmico y crasinucelado, placentación apical. El polen es tricolpado, esferoidal, suboblato, con los mesocolpos prominentes y la exina reticulada y engrosada en los bordes de los colpos (imperfectamente reticulada con muros sinuosos en Gunnera herteri).
Sus fruto son drupas carnosas o coriáceas, verdosa (de 0,6-2 mm) o rojiza (de hasta 8 mm), raramente blanca o amarilla, oval a globosa. Posee una semilla por fruto, con testa membranácea y endospermo abundante, oleoso, con granos de almidón, aleurona y cristaloides, embrión muy pequeño, excéntrico, apical, con 2 cotiledones, endospermo celular.
Su número cromosómico es 2n=34; x=11, 12, 17, 18.
Ecología
[editar]
Las flores son anemófilas y las hermafroditas, protándricas. Las especies con drupas coloreadas presentan una dispersión ornitócora. Crecen en hábitats frescos o fríos, húmedos o anegados (helófitas), como pantanos, bordes de ríos, cascadas y cerca de pozos, desde el nivel del mar a altitudes superiores a los 3000 m, si bien prefieren altitudes elevadas y áreas abiertas y luminosas, siendo pioneras en muchos casos. Presentan una amplia tolerancia térmica y una ruta fotosintética del tipo C3.
Simbiosis
[editar]Las plantas del género Gunnera forman una simbiosis de manera natural con cianobacterias fijadoras de nitrógeno atmosférico (N2), al parecer siempre con diferentes cepas de Nostoc punctiforme. Esta simbiosis presenta unas características particulares que la hacen diferente a otras asociaciones del mismo tipo: es la única que implica a una angiosperma, la cianobacteria se recluye en unos órganos glandulares especializados situados en el tallo, donde penetra en las células del huésped (simbiosis intracelular, un tipo de endosimbiosis) y deja de fotosintetizar, suministrando la cianobacteria nitrógeno a la planta a cambio de compuestos carbonados. La cianobacteria, que proviene del medio, penetra en la planta a través de raíces adventicias, que abortan y se transforman en órganos glandulares. Las células afectadas son mayores y más redondeadas de lo normal, y en su interior la cianobacteria desarrolla la mayor concentración registrada de heterocistes en una cianobacteria.
La simbiosis, aunque evolutivamente muy avanzada por ser intracelular, es, sin embargo, facultativa, ambos simbiontes sobreviven separados y pueden reconstituir su asociación si se les vuelve a poner en contacto; es menos interdependiente que la existente entre otras angiospermas y sus simbiontes como Rhizobium y también menos coordinada: las glándulas se forman incluso si la cianobacteria está ausente, lo cual implica que contenerlos no es su misión principal, ya que el conjunto de genes involucrado en la formación de las glándulas inicia su actividad sin que ésta la desencadene el huésped. Las glándulas están formadas por 6-9 papilas, de las que 5-8 son periféricas y algo arqueadas hacia fuera y una papila central es más grande, larga y erecta. Las papilas se forman en el interior del tallo desde el momento de la germinación de la plántula, por debajo de cada inserción peciolar, crecen hacia la superficie externa y atraviesan la epidermis, apareciendo externamente, mientras que en su interior se forman unos canales entre las papilas.
Las glándulas producen mucílago que fluye por los canales al exterior. Este mucílago es rico en carbohidratos, ácidos tánicos y otros polifenoles de acción antimicrobiana, es de carácter ácido y parece contener además sustancias antifúngicas. A medida que la planta crece, se forman glándulas bajo las hojas nuevas. Infectadas o no, las glándulas cesan en un momento dado de producir mucílago y de crecer, se forma una capa meristemática externa que produce hacia fuera unas capas pardas que dan a la glándula un aspecto de corcho. A partir de este momento, la entrada de cianobacterias no es posible, los canales y las cavidades desaparecen y el tejido glandular se congestiona y empieza a desintegrarse. La cianobacteria alcanza el mucílago bien por acinetos, bien por hormogonios, atraídos al parecer quimiotácticamente por los compuestos fenólicos presentes en él y a continuación empieza a dividirse repetidamente, produciendo hormogonios, se introduce en la glándula y penetra en el interior de unas células del meristemo glandular con paredes finas. Las paredes celulares se disuelven y las células se hinchan al contacto con el simbionte. El proceso de acceso al interior de la célula del hospedante no está claro, pero los hormogonios quedan incluidos dentro de una cavidad de membrana plasmática dentro de la célula, donde los filamentos hormogoniales revierten su diferenciación para dar lugar a filamentos con células vegetativas y heterocistes, siendo éstos los que fijan el N2 de manera anaerobia, mediante una nitrogenasa. Los filamentos de la cianobacteria se reparten entre las células hijas resultantes de la mitosis de una célula simbiótica. El tejido glandular contiene además células sin filamentos, menores y más alargadas, formando estructuras ramificadas entre las células con filamentos. Estas células tienen numerosos amiloplastos, lo que indica que pueden tener un papel en el intercambio de metabolitos con la cianobacteria, fundamentalmente de glúcidos. El tamaño superficial de las glándulas varía con cada especie de Gunnera, entre 2 y 20 mm de diámetro, pero es típico de cada una. La eficiencia de la fijación de N2 es muy alta, llegando a alcanzar en algunas especies (Gunnera arenaria) los 72 kg ha-1 año-1, siendo menor en las especies más grandes, que cuentan con un menor número de glándulas por unidad de área; la fijación es mayor en las partes jóvenes de la planta, por debajo del meristemo apical y decrece hacia la base de la misma. El nitrógeno se transfiere al hospedante en forma de un aminoácido de identidad desconocida por el momento. Este aporte extraordinario de nitrógeno permite el rápido crecimiento de las plantas de Gunnera, que muestran una baja capacidad de captación de los nitratos del suelo.
En el mucílago producido por las glándulas repartidas por la planta se han encontrado asimismo bacterias, hongos (Fusarium, Penicillium, Alternaria), algas y animales microscópicos, pero solo un alga verde se ha encontrado viviendo intracelularmente en las mismas.
Fitoquímica
[editar]
Los rizomas de Gunnera tinctoria contienen un 9,3% de taninos y las hojas de Gunnera manicata contienen una alta concentración de un elagitanino. Gunnera perpensa contiene el alcaloide celastrina. No cianogenéticas, no se han encontrado iridoides ni proantocianidinas, pero sí kaempferol, quercetina, flavonoles y ácido elágico.
Usos
[editar]Los peciolos de Gunnera tinctoria (nalcas) se comen en el sur de Chile y de Argentina, en fresco en ensaladas, pero también en mermelada o en licores, y los tallos (depes) tienen propiedades medicinales; sus hojas (pangues) se usan para cubrir el curanto y la mella (platos tradicionales de Chiloé); sus tallos y rizomas se usan localmente para el curtido y el tinte (en negro) de tejidos.
La decocción del rizoma de Gunnera perpensa se usa en medicina tradicional en el sur de África para facilitar el parto o como abortivo por su marcada acción sobre la musculatura uterina. Gunnera manicata es apreciada como planta ornamental, pero debido a su capacidad para escaparse y aclimatarse, compitiendo ventajosamente con especies locales, tiene en muchos países la consideración de planta invasora (p. ej., en Australia y Nueva Zelanda). Lo mismo ocurre con Gunnera tinctoria en Gran Bretaña e Irlanda.
La visita a las especies gigantes del subgénero Panke en las islas Hawái es una atracción turística. Las hojas de Gunnera insignis se usan en Costa Rica como sombrillas improvisadas.
Posición sistemática
[editar]Las gunneráceas son un grupo de Angiospermas que se incluyen en el clado Eudicotiledóneas. Se han relacionado usualmente con las haloragáceas de manera tentativa. Basándose en datos moleculares y morfológicos, el APW (Angiosperm Phylogeny Website)[2] considera que forma parte del Orden Gunnerales, siendo el grupo hermano de las mirotamnáceas en el grado basal de las Eudicotiledóneas (cf. AP-website).
Tradicionalmente el género Gunnera se venía considerando como integrante de la familia de las haloragáceas, a falta de otro sitio mejor, basándose en las flores simples, epíginas y dímeras y su apetencia por hábitats húmedos, aunque fuesen muy diferentes en otros aspectos. En sistemas posteriores, se colocó en su propia familia, las gunneráceas, junto a las haloragáceas, siempre dentro del orden Myrtales. Dahlgren (1975)[3] movió la familia a las cercanías de las Saxifragaceae, mientras que Takhtajan (1980)[4] colocó las gunneráceas en un orden Gunnerales dentro de las Saxifraganae y, por su parte, Cronquist (1981)[5] las colocó, junto con las haloragáceas en un orden Haloragales dentro de las rósidas, pero solo el advenimiento de los análisis filogenéticos moleculares permitió dilucidar la cuestión, emplazándola como una línea basal aislada del clado de las rósidas-cariofílidas-astéridas y hermana de las mirotamnáceas.
Los análisis de las secuencias de genes cloroplastidiales y nucleares han permitido resolver con bastante nitidez la filogenia interna del género. La especie basal y hermana de todas las demás especies perennes es la especie anual sudamericana Gunnera herteri. La siguiente rama la forma la especie africana G. perpensa, que es la hermana de dos clados bien definidos: uno formado por la especie asiática G. macrophylla como hermana de las especies que forman el subgénero Milligania, y otro clado, hermano del anterior, con dos grupos hermanos, los subgéneros Misandra y Panke. Dentro de este último, las dos especies hawaianas forman un grupo monofilético hermano de las especies americanas. El siguiente dendrograma ilustra las relaciones expuestas:
| Gunnerales |
| ||||||||||||||||||||||||||||||||||||
Gunnera presenta una distribución austral disyunta de tipo relíctico, similar a la de otros géneros de plantas, como Nothofagus, y animales. Se suele interpretar como el resultado de la división del antiguo continente de Gondwana, iniciada durante el Jurásico inferior, hace unos 180 ma. La existencia del género está atestiguada por pólenes fósiles (Tricolpites reticulatus) que se le atribuyen, del Turoniense (Cretácico superior, hace unos 90 ma) de Sudamérica. La divergencia estimada de Gunnera y Myrothamnus se ha establecido en los 115-118 ma, lo que coincide bastante bien con el inicio de la separación de África y Sudamérica hace unos 127-137 ma.
El único género Gunnera L., 1767 tiene como especie tipo Gunnera perpensa L., 1767. Está dedicado a Johan Ernst Gunnerus. Se divide en seis subgéneros, confirmados mediante secuenciación de ADN, a saber:
- * Subgénero Ostenigunnera Mattf., 1933. Especie tipo: Gunnera herteri Osten, 1932.
- Hierbas monoicas, anuales, menudas (2-4 cm), glabras, con tallos ramificados, delgados, con largos entrenudos. Limbos foliares hasta de 14 mm, flabeliformes, hasta con 20 lóbulos dentiformes terminados en un hidatodo, vaina foliar con lóbulos dentiformes. Sin brácteas, estolones o rizomas. Con 2-5 coléteres axilares inconspicuos. Nudos unilacunares. Inflorescencias interaxilares. Racimos de unos 10 mm de largo, con 2-7 flores masculinas en la mitad apical (frecuentemente reducidas a una antera) y las femeninas en la mitad basal (sin perianto).
- * Especie Gunnera herteri Osten, 1932, en charcas freáticas entre las dunas costeras del sur de Brasil, donde se llama urtiguinha das dunas y Uruguay. Especie en peligro.
- * Subgénero Gunnera s. str.[6]

- Hierbas de tamaño medio, con rizoma horizontal, ramificado periódicamente, de 10-20 mm de grueso. Sin brácteas ni estolones. Hojas de peciolo largo y limbo cordiforme o reniforme hasta de 28 cm de ancho, con el borde densamente dentado-crenado. Partes jóvenes pubescentes. Inflorescencia tirsoide, hasta de 40 cm, con flores hermafroditas.
- * Especie Gunnera perpensa L., 1767,[7] conocida de Sudáfrica, Zimbabue, Mozambique, República Democrática del Congo, Ruanda, Burundi, Kenia, Tanzania, Sudán, Etiopía y Madagascar.
- * Subgénero Pseudogunnera (Oerst., 1857) Schindl., 1905.[8]
- Hierbas monoicas, de tamaño medio, con largos estolones y tallos erectos, cortos, de 10-20 mm de diámetro, los estolones con brácteas pareadas opuestas. Hojas moderadamente grandes, hasta de 50 cm de ancho, envainantes en la base, con peciolo hasta de 1 m y limbo reniforme a cordiforme, irregularmente lobulado, los bordes irregularmente dentados y oscurecidos, con nervadura prominente en el envés y vesículas en el haz. Inflorescencias hasta de 50 cm, las flores superiores masculinas con sépalos y pétalos, las inferiores femeninas solo con sépalos.
- * Especie Gunnera macrophylla Blume, 1825-1826,[9] extendida desde Java, Sumatra y las Filipinas hasta Nueva Guinea y las Islas Salomón. Probablemente se trata de un complejo de especies cercanas.
- * Subgénero Milligania (Hook.f., 1840) Schindl., 1905.[10] Especie tipo: Milligania cordifolia Hook.f., 1840.

- Hierbas pequeñas, estoloníferas, cespitosas, con tallos erectos cortos que llevan una roseta de hojas, sin escamas, con un par de brácteas en el extremo de los estolones. Hojas con peciolo hasta de 20 mm, el limbo orbicular-reniforme, subcordiforme, ovado o elíptico, hasta de 5 cm de largo. Inflorescencias separadas para cada sexo en la misma planta (monoecia), excepto en Gunnera monoica, con racimos de flores masculinas en la parte apical y femeninas en la basal. Racimos de 1-5 cm de largo, flores masculinas con perianto completo, las femeninas solo con sépalos. Seis especies, una de Tasmania y cinco de Nueva Zelanda.
- * Especie Gunnera cordifolia (Hook.f., 1840) Hook.f., 1860,[11] de Tasmania.
- * Especie Gunnera densiflora Hook.f., 1864
- * Especie Gunnera dentata Kirk, 1895[12]
- * Especie Gunnera hamiltonii Kirk ex W.S.Ham., 1885
- * Especie Gunnera monoica Raoul, 1844[13]
- * Especie Gunnera prorepens Hook.f., 1852[14]
- * Subgénero Misandra (Comm. ex Juss., 1789) Schindl., 1905.[15] Especie tipo: Gunnera magellanica Lam., 1789.
.jpg)
- Hierbas estoloníferas pequeñas, dioicas. Tallos erectos cortos con hojas alternando con escamas en forma de ócrea, el extremo del estolón con una ócrea cónica cubriendo la yema apical. Hojas con peciolo de 2-25 cm de largo y limbo reniforme o reniforme-orbicular hasta de 11 cm de ancho, escasamente lobulado. Inflorescencias hasta de 15 cm, todas las flores sin pétalos. Tres especies, desde la Tierra del Fuego y las islas Malvinas hasta Colombia.
- * Especie Gunnera lobata Hook.f., 1846, de Argentina y Chile
- * Especie Gunnera magellanica Lam., 1789,[16] de Argentina, Bolivia, Chile, Colombia, Ecuador, Malvinas y Perú, con hojas de 5 a 9 cm de ancho sobre tallos de 8 a 15 cm de largo.
- * Especie Gunnera reichei Schindl., 1905, de Chile
- * Subgénero Panke (Molina, 1782) Schindl., 1905.[17] Especie tipo: Gunnera petaloidea Gaudich., 1830.
- Hierbas perennes grandes a gigantes (hasta de 6 m de altura), de tallos erectos, engrosados y carnosos, hasta de 3 m de altura y 40 cm de diámetro, a menudo decumbentes con la edad, rara vez ramificados, partes jóvenes y yema terminal cubiertas de numerosas escamas linear-lanceoladas, enteras o laciniadas, hasta de 40 cm de largo, frecuentemente de un color naranja brillante. Hojas con peciolo hasta de 2,7 m de largo y limbos usualmente palmatilobados, hasta de 4 m de diámetro o más. Inflorescencias hasta de 50 cm en espiga o racimo compuestos, flores masculinas con perianto completo, las femeninas solo con sépalos. Consta de 50 especies, repartidas por México, Centroamérica y Sudamérica, y los archipiélagos de Juan Fernández y Hawái. Las especies gigantes están consideradas entre las hierbas más grandes del planeta.
- * Especie Gunnera aequatoriensis L.E.Mora, 1978, de Ecuador, especie considerada "Vulnerable D2" por la IUCN[18]
- * Especie Gunnera annae Schindl., 1905, de Bolivia y Perú, denominada quirisulla
- * Especie Gunnera antioquensis L.E.Mora, 1978, de Colombia
- * Especie Gunnera apiculata Schindl., 1905,[19] de Bolivia y Argentina
- * Especie Gunnera atropurpurea L.E.Mora, 1978,[20] de Colombia
- * Especie Gunnera berteroi Phil., 1872,[21] de Bolivia y Chile
- * Especie Gunnera bogotana L.E.Mora, 1978, de Colombia
- * Especie Gunnera bolivari J.F.Macbr., 1931, de Ecuador y Perú
- * Especie Gunnera boliviana Morong ex Rusby, 1896, de Bolivia
- * Especie Gunnera bracteata Steud. ex Benn., 1838,[22] del archipiélago Juan Fernández
- * Especie Gunnera brephogea Linden & André, 1872,[23] de Colombia y Ecuador
- * Especie Gunnera caucana L.E.Mora, 1984, de Colombia
- * Especie Gunnera colombiana L.E.Mora, 1978, de Colombia y Ecuador
- * Especie Gunnera commutata Blume, 1856, de Chile
- * Especie Gunnera cuatrecasasii L.E.Mora, 1984, de Colombia
- * Especie Gunnera diazii L.E.Mora, 1984, de Colombia
- * Especie Gunnera ecuadoriana Gilli, 1981, de Ecuador
- * Especie Gunnera garciae-barrigae L.E.Mora, 1984, de Colombia
- * Especie Gunnera hernandezii L.E.Mora, 1978, de Colombia
- * Especie Gunnera insignis (Oerst., 1857) A.DC., 1868,[24] de México, Nicaragua, Costa Rica, Panamá y Colombia, llamada sombrilla de pobre


- * Especie Gunnera kauaiensis Rock, 1930,[25] de las islas Hawái
- * Especie Gunnera killipiana Lundell, 1940, de México
- * Especie Gunnera lozanii L.E.Mora, 1984, de Colombia
- * Especie Gunnera magnifica H.St.John, 1957, de los Andes colombianos presenta las yemas foliares más grandes que en cualquier otra planta, hasta de 60 cm de largo y 40 cm de grosor. Tiene las hojas gruesas con peciolos de 2,7 m de altura. La inflorescencia gruesa de flores pequeñas y rojizas alcanza 2,3 m de altura y pesa aproximadamente 13 kg.
- * Especie Gunnera manicata Linden ex Delchev., 1867,[26] natural de Serra do Mar, montañas del sudeste de Brasil, es quizás la especie más grande, con hojas de alrededor de 1,5 m a 2 m de ancho, pero excepcionalmente (como en Narrow-water Park, County Cork, Irlanda, en 1903) hasta 3,4 m de ancho, sostenida por gruesos rizomas y suculentos peciolos de hasta 2,5 m de largo. Germina en condiciones muy húmedas (no lluviosas) con temperaturas de 24 a 29 °C. Es muy apreciada como ornamental.
- * Especie Gunnera margaretae Schindl., 1911, de Bolivia y Perú
- * Especie Gunnera masafuerae Skottsb., 1914, del archipiélago Juan Fernández. Puede tener hojas hasta de 2,9 m de ancho, sobre fuertes peciolos de 1,5 m de largo y 11 cm de grosor.
- * Especie Gunnera mexicana Brandegee, 1922, de México, Guatemala y Honduras
- * Especie Gunnera morae L.Wanntorp & Klack., 2006, de Colombia
- * Especie Gunnera peltata Phil., 1856,[27] del archipiélago Juan Fernández, con frecuencia tiene un tronco derecho de 5,5 m de altura y 25-30 cm de grosor, y hojas gruesas, que tienen hasta 2 m de ancho.
- * Especie Gunnera peruviana J.F.Macbr., 1931, de Bolivia y Perú
- * Especie Gunnera petaloidea Gaudich., 1830,[28] de las islas Hawái
- * Especie Gunnera pilosa Kunth, 1817, de Colombia, Ecuador y Bolivia
- * Especie Gunnera pittierana V.M.Badillo & Steyerm., 1973, de Venezuela, popularmente llamada paraguas de pobre, es endémica del Parque Nacional Henri Pittier y es la planta herbácea con mayor tamaño de hoja. En este parque y a altitudes de los 850 a 1500 m se las ha encontrado con tamaños que superan los 4 m de ancho.
- * Especie Gunnera quitoensis L.E.Mora, 1978, de Ecuador
- * Especie Gunnera rheifolia Schindl., 1905, de Perú
- * Especie Gunnera saint-johnii (L.E.Mora, 1978) L.E.Mora, 1984,[29] de Colombia
- * Especie Gunnera sanctae-marthae L.E.Mora, 1984, de Colombia
- * Especie Gunnera schindleri L.E.Mora, 1978, de Argentina y Bolivia
- * Especie Gunnera schultesii L.E.Mora, 1984, de Colombia
- * Especie Gunnera silvioana L.E.Mora, 1984, de Colombia
- * Especie Gunnera steyermarkii L.E.Mora, 1978, de Venezuela
- * Especie Gunnera tacueyana L.E.Mora, 1984, de Colombia
- * Especie Gunnera tajumbina L.E.Mora, 1984, de Colombia
- * Especie Gunnera talamancana H.Weber & L.E.Mora, 1958, de Costa Rica y Panamá
- * Especie Gunnera tamanensis L.E.Mora, 1984, de Colombia
- * Especie Gunnera tayrona L.E.Mora, 1978, de Colombia
- * Especie Gunnera tinctoria (Molina, 1782) Mirb., 1805,[30] de Argentina y Chile, introducida en Gran Bretaña, Irlanda y Oregón
- * Especie Gunnera venezolana L.E.Mora, 1978, de Venezuela. Está dividida en dos subespecies:
- * subsp. venezolana, que es la forma típica
- * subsp. tachirensis L.E.Mora, 1978
- * Especie Gunnera vestita Schindl., 1905, de Chile.
Híbridos
[editar].jpg)
Las especies de Gunnera muestran una moderada tendencia al cruzamiento interespecífico. Se conocen los siguientes híbridos:
- Notoespecie Gunnera × katherine-wilsoniae L.D.Gómez, 1983 (Gunnera talamancana × Gunnera insignis), de Costa Rica
- Notoespecie Gunnera × strigosa (Kirk, 1895) Colenso, 1883 (Gunnera monoica × Gunnera dentata), de Nueva Zelanda
- Notoespecie Gunnera × mixta Kirk, 1895 (Gunnera monoica × Gunnera prorepens), de Nueva Zelanda
- Gunnera peltata × Gunnera bracteata, en la isla de Robinsón Crusoe (Archipiélago Juan Fernández).
Fósiles
[editar]Aparte del polen fósil Tricolpites reticulatus ya mencionado, se conoce polen fósil de Gunnera de estratos campanienses (Cretáceo Superior) de Australia, Nueva Zelanda, África y la Antártida, probablemente derivados de los ancestros de los subgéneros actuales. Asimismo, se han hallado tanto pólenes fósiles como macrofósiles de hojas en estratos del Cretáceo Superior de Norteamérica, lo que podría implicar que los ancestros del subgénero Panke colonizaron Sudamérica y las islas Hawái desde allí, para desaparecer después, lo que concuerda con la posición basal de las especies hawaianas en el subgénero. En las islas Hawái el polen de Gunnera solo se conoce desde el Pleistoceno, lo que indica una colonización más reciente por dispersión a larga distancia. Es evidente de los datos paleopolínicos que el género ha desaparecido de áreas del hemisferio norte donde estuvo presente durante el Cretáceo Superior y los inicios del Cenozoico. Asimismo, la distribución de Gunnera macrophylla debe atribuirse a una dispersión hacia el norte de un componente de una biota gondwánica relacionado con las formas neozelandesas. Todos estos hechos concuerdan con su consideración actual como una de las familias más primitivas de las dicotiledóneas nucleares o eudicotiledóneas (también llamadas Tricolpadas).
Notas
[editar]Las estructuras escamosas que crecen en el tallo de las especies de los subgéneros Misandra y Panke se han interpretado de muy diversas maneras según los autores: lígulas, estípulas intrapeciolares, escamas u hojas no desarrolladas, lo que supondría, en este último caso, la existencia de heterofilia. La cuestión no está aún resuelta.
A veces es posible encontrar flores anómalas. En Gunnera tinctoria se encuentran flores con un ovario bilocular y, por tanto, con dos óvulos.
Referencias
[editar]- ↑ Fernández Herrera, D. (1984). «Estudio anatómico y morfológico de Gunnera insignis (Oerst.) A. DC.». Revista de Biología Tropical 32 (2). p. 197-202.
- ↑ Stevens, P (2013). «Angiosperm Phylogeny Website. Version 12, July 2012.». [En línea].
- ↑ Dahlgren, R. (1975). «A system of classification of the angiosperms to be used to demonstrate the distribution of characters.». Botaniska Notiser 128. p. 119-147.
- ↑ Takhtajan A. (1980). «Outline of the classification of flowering plants (Magnoliophyta).». Botanical Review 46. p. 225-359.
- ↑ Cronquist A. (1981). An Integrated System of Classification of Flowering Plants. Columbia University Press, New York. ISBN 0-231-03880-1.
- ↑ Sinónimo: Perpensum Burm.f., 1768.
- ↑ Sinónimos: Perpensum blitispermum Burm.f., 1768; Gunnera calthifolia Salisb., 1796; Gunnera perpensa var. alpina T.C.E.Fr., 1924; Gunnera perpensa var. angusta Schindl., 1905; Gunnera perpensa var. kilimandscharica Schindl., 1905
- ↑ Sinónimo: Pseudo-Gunnera Oerst., 1857
- ↑ Sinónimo: Gunnera erosa Blume, 1825; Gunnera reniformis Ridl., 1916
- ↑ Sinónimo: Milligania Hook.f., 1840; Misandropsis Oerst., 1857
- ↑ Sinónimo: Milligania cordifolia Hook.f., 1840
- ↑ Sinónimos: Gunnera arenaria Cheeseman ex Kirk, 1895; Gunnera densiflora var. depressa Kirk, 1895
- ↑ Sinónimos: Gunnera microcarpa Kirk, 1895; Gunnera ovata Petrie, 1893 ; Gunnera monoica var. albocarpa Kirk, 1895; Gunnera monoica var. ramulosa Kirk, 1895
- ↑ Sinónimo: Gunnera flavida Colenso, 1886
- ↑ Sinónimo: Misandra Comm.. ex Juss.., 1789
- ↑ Sinónimos: Gunnera plicata Vahl, 1804; Gunnera falklandica Hook., 1842; Gunnera integrifolia Blume, 1856; Gunnera reniformis Gay ex Blume, 1856
- ↑ Sinónimos: Panke Molina, 1782; Gunneropsis Oerst., 1857; Pankea Oerst., 1857
- ↑ «The IUCN Red List of threatened species». Consultado el 1 de abril de 2013.
- ↑ Sinónimo: Gunnera chilensis A.DC., 1868, nom. illeg.
- ↑ Variedades: var. atropurpurea; var. caquetana L.E.Mora, 1984; var. chocoana L.E.Mora, 1984; var. munchicana L.E.Mora, 1984; var. sibundoya L.E.Mora, 1984
- ↑ Sinónimo: Gunnera laxiflora Phil., 1894
- ↑ Sinónimo: Gunnera insularis Phil., 1856; Gunnera scabra Phil., 1856, nom. illeg.; Gunnera pyramidalis Schindl., 1905
- ↑ Sinónimos: Gunnera brephogea subsp. magna L.E.Mora, 1978
- ↑ Sinónimo: Pankea insignis Oerst., 1857; Gunnera insignis fo. albovariegata L.D.Gómez, 1983; Gunnera wendlandii Reinke ex Schindl., 1905
- ↑ Sinónimo: Gunnera dominii Krajina, 1930
- ↑ Sinónimo: Gunnera brasiliensis Schindl., 1925
- ↑ Sinónimo: Gunnera glabra Phil.,, 1856
- ↑ Sinónimos: Gunnera petaloidea var. kaalaensis Krajina, 1930; Gunnera petaloidea var. mauiensis Krajina, 1930; Gunnera eastwoodiae H.St.John, 1946; Gunnera makahaensis H.St.John, 1946; Gunnera molokaiensis H.St.John, 1946
- ↑ Sinónimo: Gunnera pilosa subsp. saint-johnii L.E.Mora, 1978
- ↑ Sinónimos: Panke tinctoria Molina, 1782; Panke acaulis Molina, 1782; Gunnera chilensis Lam., 1789; Panke caulescens J.F.Gmel., 1791; Gunnera scabra Ruiz & Pav., 1798. Variedades: Gunnera chilensis var. meyerii L.E.Mora, 1978; Gunnera chilensis var. valdiviensis L.E.Mora, 1978.
Bibliografía
[editar].jpg)
- Bergman, B., Johansson, C., Söderbäck, E. (1992). «Tansley Review No. 42. The Nostoc-Gunnera Symbiosis.». New Phytologist 122 (3). p. 379-400.
- Rutishauser, R., Wanntop, L., Pfeifer, E. (2004). «´Gunnera herteri - developmental morphology of a dwarf from Uruguay and S Brazil (Gunneraceae).». Plant Systematics and Evolution 248. p. 219-241.
- Wanntorp, L., Wanntorp, H.-E. & Källersjö M. (2002). «Phylogenetic Relationships of Gunnera based on Nuclear Ribosomal DNA ITS Region, rbcL and rps16 Intron Sequences». Systematic Botany 27 (3). p. 512-521.
- Wanntorp, L., Wanntorp, H.-E., Oxelman, B. & Källersjö M. (2001). «Phylogeny of Gunnera». Plant Systematics and Evolution 226. p. 85-107.
- Watson, L., and Dallwitz, M.J. (1992 onwards). «The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 19th December 2012.». Archivado desde el original el 3 de enero de 2007. Consultado el 15 de marzo de 2013.
- Watson, L., and Dallwitz, M.J. (1992 onwards). «Landcare Research Manaaki Whenua». Consultado el 10 de marzo de 2013.
- Wilkinson, H. P., Wanntorp, L. (2007). «Gunneraceae.». Kubitzki, K. (Editor). The Families and Genera of Vascular Plants. IX. Flowering Plants. Eudicots: Berberidopsidales, Buxales, Crossosomatales, Fabales p.p., Geraniales, Gunnerales, Myrtales p.p., Proteales, Saxifragales, Vitales, Zygophyllales, Clusiaceae Alliance, Passifloraceae Alliance, Dilleniaceae, Huaceae, Picramniaceae, Sabiaceae. Springer-Verlag: Berlín. ISBN 3-540-32214-0.
Enlaces externos
[editar] Wikimedia Commons alberga una categoría multimedia sobre Gunneraceae.
Wikimedia Commons alberga una categoría multimedia sobre Gunneraceae. Wikispecies tiene un artículo sobre Gunneraceae.
Wikispecies tiene un artículo sobre Gunneraceae.
{kind=link}
Otro mapa con los subgéneros Archivado el 27 de mayo de 2016 en Wayback Machine.
{kind=link}
{kind=link}
Hoja gigante (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Órganos glandulares con cianobacterias en Gunnera pilosa
{kind=link}
Glándula con Nostoc simbióticos
{kind=link}
Corte de las glándulas de Nostoc mostrando los filamentos cianobacteriales intracelulares observado al microscopio Archivado el 5 de marzo de 2016 en Wayback Machine.
{kind=link}
{kind=link}
| Control de autoridades |
|
|---|
 Datos: Q2338324
Datos: Q2338324- Multimedia: Gunneraceae / Q2338324
- Especies: Gunneraceae