Tái tổ hợp di truyền
.png)
Tái tổ hợp gen là hiện tượng sinh vật bố mẹ sinh ra con có những tính trạng khác với bố mẹ do sự tổ hợp lại những gen vốn có của bố mẹ, phát sinh ra bởi sự chuyển đổi lô-cut của các gen trong hệ gen.[1], [2], [3]
Các tên khác: tái tổ hợp di truyền (genetic recobination) hoặc tái tổ hợp sinh học (biological recombination) hay tái tổ hợp DNA (DNA recombination) cũng có nội hàm này. Khi cần gọi tắt, người ta dùng tên tái tổ hợp.
Hiện nay có thể phân biệt hai kiểu chính của tái tổ hợp là: tái tổ hợp tương đồng và tái tổ hợp không tương đồng (hình 1).
Thomas Hunt Morgan được công nhận là nhà khoa học đầu tiên đã phát hiện ra hiện tượng này và giải thích hợp lí một phần bản chất của tái tổ hợp từ đầu những năm 1900. Hiện tượng mà ông phát hiện, ngày nay gọi là tái tổ hợp tương đồng.[4] Nhưng sau đó, nhiều nhà khoa học khác có nhiều phát hiện khác, bổ sung và tìm hiểu sâu hơn đến cấp độ phân tử của hiện tượng này (chi tiết hơn xem ở trang Gen hoán vị).
Lược sử khái niệm
[sửa | sửa mã nguồn]Nội dung ban đầu
[sửa | sửa mã nguồn]- Thuật ngữ tái tổ hợp gen có xuất xứ từ tiếng Anh: genetic recombination (phát âm quốc tế: /ʤɪnɛ́tɪk rɪ́jkɔmbɪnɛ́jʃən/; tiếng Việt: gi-nê-tic ri-côm-bi-nêy-sân), do Thomas Hunt Morgan đề xuất vào khoảng những năm 1900 – 1910 (xem hình 2). Trong các ngôn ngữ khác: tiếng Pháp là recombinaison génétique, tiếng Đức là Rekombination genetik v.v đều có nội hàm tương tự như trên.[5], [6], [7]
- Morgan dùng khái niệm genetic recombination (tái tổ hợp gen), sau khi ông nghiên cứu về gen liên kết và gen hoán vị ở ruồi giấm (Drosophila melanogaster) để phân biệt với khái niệm "biến dị tổ hợp" dùng để mô tả các biến dị phát sinh do phân li độc lập các gen theo quy luật Mendel phát hiện cách lúc đó không lâu. Trong khái niệm này có từ genetic dùng để phân biệt với sự tái tổ hợp gặp ở các lĩnh vực khoa học khác như trong toán học, vật lý học...
- Một trong những thí nghiệm của Morgan liên quan đến hiện tượng này là: ruồi giấm vốn chỉ có kiểu hình màu xám, cánh dài (BV//BV) và dạng màu đen, cánh cụt (bv//bv). Nhưng trong thí nghiệm khác, thì ông đã lại thu được dạng xám, cụt (Bv//bv) và dạng đen, dài (bV//bv); nghĩa là ruồi con có kiểu hình khác bố mẹ, nhưng các tính trạng này vốn bố mẹ đã có, mà các gen quy định chúng đã đổi chỗ cho nhau do trao đổi chéo gây ra sự tổ hợp lại. Bởi vậy mới gọi là tái tổ hợp gen (gen tổ hợp lại).[8], [9]
- Xem chi tiết đã trình bày ở trang gen liên kết và trang gen hoán vị.
Nội dung mở rộng
[sửa | sửa mã nguồn]Tái tổ hợp DNA
[sửa | sửa mã nguồn]Sau đó nhiều năm, khi khái niệm gen được khoa học định nghĩa chính xác hơn và nhất là bản chất gen được khám phá nhờ Francis Crick và James Watson, thì người ta hiểu được thực chất của quá trình mà Morgan khám phá là có sự trao đổi các đoạn DNA, nên thường gọi là tái tổ hợp gen hoặc đôi khi gọi là tái tổ hợp DNA (DNA recombination).[5]
DNA tái tổ hợp
[sửa | sửa mã nguồn]Khi Di truyền học vi khuẩn phát triển mạnh, nhất là kĩ thuật chuyển gen ra đời và phát triển nhanh chóng, khái niệm recombination gene và recombination DNA thêm nội dung mới: nó dùng để chỉ gen hoặc DNA đã được tổ hợp lại một cách nhân tạo hoặc tự nhiên. Chẳng hạn: Plasmid của trực khuẩn lị E. coli đã được "ghép" gen insulin của người được gọi là DNA tái tổ hợp.
Tái tổ hợp không tương đồng
[sửa | sửa mã nguồn]- Sau khi Barbara McClintock được giải Nobel 1983 về nhiều công trình nghiên cứu của bà từ năm 1948, trong đó nổi bật là công trình về gen nhảy, thì học thuyết di truyền nhiễm sắc thể phát triển hẳn sang giai đoạn mới, đồng thời khái niệm mà chúng ta đang xét cũng mang thêm nội dung mới.
- Mô tả theo cách đơn giản nhất khái niệm gen nhảy này như sau:
- Một gen có thể thay đổi vị trí của nó bằng cách "nhảy" sang lô-cut gen khác, không có gen nào khác nhảy vào chỗ của nó.
- Gen nhảy có thể chuyển sang nhiễm sắc thể không tương đồng (hình 3). Nghĩa là không có "trao đi, đổi lại" vật chất di truyền.
- Bởi thế, trong Di truyền học đã phát sinh thêm thuật ngữ "tái tổ hợp không tương đồng", để phân biệt với sự tổ hợp lại các gen do gen hoán vị, sau này gọi lại là tái tổ hợp tương đồng mà Morgan phát hiện.
- Xem chi tiết vấn đề này ở trang gen nhảy.
-
 Hình 2: Ảnh chụp lại trang sách của Morgan (1916) mô tả cơ chế tái tổ hợp gen đầu tiên.
Hình 2: Ảnh chụp lại trang sách của Morgan (1916) mô tả cơ chế tái tổ hợp gen đầu tiên. -
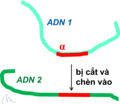 Hình 3: Gen anpha "nhảy" từ DNA 1 sang DNA 2.
Hình 3: Gen anpha "nhảy" từ DNA 1 sang DNA 2.

.png)
Đặc điểm chính
[sửa | sửa mã nguồn]- Trong tái tổ hợp có sự sắp xếp lại và thậm chí cấu trúc lại một phần hoặc cả hệ gen (genome) của tế bào sống.
- Có sự chuyển vị (translocation) của gen, thường là chuyển đổi vị trí một vài cụm gen hoặc cả một hay nhiều đoạn nhiễm sắc thể với nhiều gen.
- Do đó, thường dẫn đến thay đổi lô-cut gen, thậm chí có phân tử DNA bị mất gen, trong khi có DNA khác lại được nhận thêm gen.[10]
- Kết quả là dẫn đến phát sinh tổ hợp alen (allelic combination) mới.[11]
- Tuy ngày nay các nhà khoa học đã phát hiện ra nhiều hình thức tái tổ hợp rất khác nhau, nhưng đặc điểm cơ bản là quá trình tái tổ hợp luôn dẫn đến sự thay đổi vật chất di truyền (DNA) của tế bào hoặc cơ thể có tái tổ hợp.
Các kiểu tái tổ hợp
[sửa | sửa mã nguồn]Tái tổ hợp tương đồng
[sửa | sửa mã nguồn]- Đây là kiểu tái tổ hợp giữa hai đoạn DNA như nhau hoặc tương tự nhau, trong đó có sự trao đổi lẫn nhau.
- Kiểu này thường xảy ra tự nhiên, thường không do tác nhân đột biến gây ra (tái tổ hợp tự nhiên), hay gặp ở quá trình giảm phân của sinh vật nhân thực (nên còn gọi là tái tổ hợp giảm phân), nhưng cũng đã phát hiện ở vi khuẩn (nhân sơ).
Xem chi tiết ở trang Tái tổ hợp tương đồng.
Tái tổ hợp không tương đồng
[sửa | sửa mã nguồn]- Đặc trưng của kiểu này là sự tái tổ hợp giữa hai DNA hoặc hai nhiễm sắc thể không tương đồng với nhau.
- Trong đó có thể có nhiều dạng:
- Dạng có trao đổi vật chất di truyền cho nhau, gọi là trao đổi tương hỗ hay chuyển vị đối ứng (reciprocal translocations); trong đó có sự "trao đi, nhận lại". Sinh học phổ thông Việt Nam đã gọi dạng này chuyển đoạn tương hỗ giữa 2 nhiễm sắc thể không tương đồng.[12]
- Sự trao đổi vật chất di truyền không tương hỗ tức là chuyển vị không đối ứng (nonreciprocal translocations).[13] Trong dạng này, có "trao đi", nhưng không có "nhận lại", nghĩa là một DNA bị mất đoạn mà không được DNA kia "cho" lại chút nào.
- Nếu vật chất di truyền là cả một cánh nhiễm sắc thể hoặc cả một chiếc nguyên vẹn được chuyển vị không đối ứng thì gọi là chuyển vị Rô-bec-sơn (Robertsonian translocation).[14]
- Gen nhảy.
- Gen tái tổ hợp tự nhiên hoặc nhân tạo.
* * *
Tóm lại, tái tổ hợp gen là sự tổ hợp lại những gen vốn có của cơ thể sinh vật hoặc kết hợp thêm cả nguồn gen ngoại lai, phát sinh ra bởi sự chuyển đổi lô-cut của một số hoặc nhiều gen trong cả hệ gen, dẫn đến cấu trúc lại hệ gen. Tái tổ hợp rất quan trọng cho quá trình tiến hoá của Sinh giới.
Nguồn trích dẫn
[sửa | sửa mã nguồn]- ^ "DNA Is Constantly Changing through the Process of Recombination".
- ^ "Genetic recombination".
- ^ "Suzanne Clancy: Genetic Recombination © 2008 Nature Education".
- ^ "Eric R. Kandel:".
- ^ a b "Genetic recombination". Truy cập ngày 3 tháng 1 năm 2018.
- ^ https://www.futura-sciences.com/sciences/definitions/physique-recombinaison-4797/
- ^ https://www.biologie-seite.de/Biologie/Rekombination
- ^ Campbell: Sinh học - Nhà xuất bản Giáo dục, 2010
- ^ Phạm Thành Hổ: Di truyền học - Nhà xuất bản Giáo dục, 1998
- ^ "Reciprocal translocation". Bản gốc lưu trữ ngày 4 tháng 10 năm 2018. Truy cập ngày 1 tháng 11 năm 2018.
- ^ https://www.biology-online.org/dictionary/Genetic_recombination
- ^ SGK "Sinh học 12" - Nhà xuất bản Giáo dục, 2016.
- ^ "Phillip McClean: Translocations".
- ^ "Bản sao đã lưu trữ". Bản gốc lưu trữ ngày 11 tháng 9 năm 2017. Truy cập ngày 1 tháng 11 năm 2018.
Liên kết ngoài
[sửa | sửa mã nguồn]- Animations – homologous recombination: Animations showing several models of homologous recombination
- The Holliday Model of Genetic Recombination
- MeSH Genetic+recombination
- Animated guide to homologous recombination.
 GIẢM
20%
GIẢM
20%
 GIẢM
23%
GIẢM
23%
 GIẢM
11%
GIẢM
11%

 GIẢM
10%
GIẢM
10%
 GIẢM
41%
GIẢM
41%



