Polypore

Los políporos son un grupo de hongos que forman grandes cuerpos fructíferos con poros o tubos en la parte inferior (ver Delimitación para las excepciones). Son un grupo morfológico de basidiomicetos similares a las setas con láminas y los hongos hidnoides, y no todos los políporos están estrechamente relacionados entre sí. Los políporos también se denominan hongos de soporte o hongos de repisa, y se caracterizan por producir cuerpos fructíferos leñosos, con forma de repisa o de soporte, u ocasionalmente circulares, que se denominan conchas.
La mayoría de los políporos habitan en los troncos o ramas de los árboles consumiendo la madera, pero algunas especies que habitan en el suelo forman micorrizas con los árboles. Los políporos y los hongos corticioides relacionados son los agentes más importantes de la descomposición de la madera, y desempeñan un papel muy importante en el ciclo de los nutrientes y la producción de dióxido de carbono de los ecosistemas forestales.[1]
Se han descrito para la ciencia más de mil especies de políporos,[2] pero una gran parte de la diversidad sigue siendo desconocida incluso en zonas templadas relativamente bien estudiadas. Los políporos son mucho más diversos en los bosques naturales antiguos con abundante madera muerta que en los bosques más jóvenes gestionados o en las plantaciones. En consecuencia, varias especies han disminuido drásticamente y están en peligro de extinción debido a la tala y la deforestación.
Los poliporos se utilizan en la medicina tradicional y se estudian activamente por su valor medicinal y sus diversas aplicaciones industriales. Varias especies de poliporas son graves patógenos de las plantaciones de árboles y son las principales causas del deterioro de la madera.




Los conchos, los cuerpos fructíferos de las poliporas, se sitúan en una agrupación planar cercana de filas horizontales separadas o interconectadas. Los conchos pueden variar desde una sola fila de unos pocos casquillos, hasta decenas de filas de casquillos que pueden pesar varios cientos de kilos. Se encuentran principalmente en árboles (vivos y muertos) y en restos leñosos gruesos, y pueden parecerse a las setas. Algunos forman cuerpos fructíferos anuales, mientras que otros son perennes y crecen año tras año. Los hongos de corchete suelen ser resistentes y robustos y producen sus esporas, llamadas basidiosporas, dentro de los poros que suelen formar la superficie inferior.
Clasificación
[editar]Dado que los hongos de corcho se definen por su forma de crecimiento y no por su filogenia, el grupo contiene miembros de múltiples clados. Aunque el término "hongos de corchete" se reservaba clásicamente para los poliporos, los estudios moleculares han revelado algunas relaciones extrañas. El hongo del bistec, muy conocido, es en realidad un miembro de los agáricos. Otros ejemplos de hongos de abrazadera son el estante de azufre, el abedul, la silla de montar de la dríade, la concha de artista y la cola de pavo. El nombre de políporos se utiliza a menudo para un grupo que incluye muchos de los hongos duros o coriáceos, que a menudo carecen de un estipe, que crecen directamente en la madera. "Polypore" deriva de las palabras griegas poly, que significa "mucho" o "muchos", y poros, que significa "poro".[5]
El grupo incluye muchas formas diferentes que son comunes en los bosques tropicales, incluyendo los duros "hongos de copa" y los hongos "de concha", "de placa" y "de ménsula" que se encuentran comúnmente creciendo de troncos y árboles muertos aún en pie.
Descripción
[editar]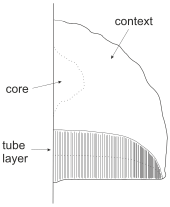
El individuo fúngico que desarrolla los cuerpos fructíferos que se identifican como poliporas reside en el suelo o en la madera como micelio. Los pólporos suelen estar restringidos a los árboles huéspedes de hoja caduca (angiospermas) o de coníferas (gimnospermas). Algunas especies dependen de un solo género de árbol (por ejemplo, Piptoporus betulinus en abedules, Perenniporia corticola en dipterocarpos).
Las formas de los cuerpos fructíferos de los poliporos van desde la forma de hongo hasta las finas manchas efusivas (costras) que se desarrollan en la madera muerta. Los cuerpos fructíferos perennes de algunas especies que crecen en árboles vivos pueden superar los 80 años (por ejemplo, Phellinus igniarius[6]). La mayoría de las especies de políporas desarrollan nuevos cuerpos fructíferos de corta duración anualmente o varias veces al año. La fructificación abundante tiene lugar durante el otoño o la temporada de lluvias.
La estructura de los cuerpos fructíferos es sencilla. Los cuerpos fructíferos efusivos o resupinados suelen estar formados por dos capas: una capa tubular de tubos dispuestos verticalmente que se abren hacia abajo, y una capa de soporte llamada subículo que sostiene y fija los tubos al sustrato. En los cuerpos fructíferos con capuchón (cuerpos fructíferos pileados) el tejido entre la superficie superior y la capa de poros se denomina contexto. Unos pocos poliporos (por ejemplo, Fomes fomentarius e Inocutis rhaedes) también tienen un núcleo entre el contexto y el sustrato. Una minoría de poliporas también tiene un tallo (estipe) que se une a la tapa lateral o centralmente, dependiendo de la especie.
Los tubos polipoides tienen una estructura similar a la de un panal, donde los tubos individuales se han fusionado. Sus lados están cubiertos por una superficie formadora de esporas, el himenio. Los tubos ofrecen refugio a las esporas en desarrollo y ayudan a aumentar el área de la superficie productora de esporas. El tamaño y la forma de los poros varían mucho entre especies, pero poco dentro de una misma especie: algunas Hexagonia spp. tienen poros de 5 mm de ancho, mientras que los poros de Antrodiella spp. son invisibles a simple vista, con 15 poros por mm. En general, cuanto más grandes son los poros, más grandes son las esporas. Algunas poliporas producen esporas asexuales (clamidosporas o conidios) en la superficie superior de su sombrero (por ejemplo, Echinopora aculeifera, Oligoporus ptychogaster) o sin la presencia de un cuerpo fructífero sexual (por ejemplo, Inonotus rickii, Heterobasidion spp.).
-
 Cuerpos frutales de Polyporus sp. con tallo (Indonesia)
Cuerpos frutales de Polyporus sp. con tallo (Indonesia) -
 Concha perenne de Fomitopsis pinicola en abeto
Concha perenne de Fomitopsis pinicola en abeto -
 Cuerpos fructíferos efusivos de Meruliopsis taxicola en un tronco de pino
Cuerpos fructíferos efusivos de Meruliopsis taxicola en un tronco de pino -
 lado inferior poliporoso con poros/tubos claramente visibles
lado inferior poliporoso con poros/tubos claramente visibles




Ecología
[editar]Los hongos de sujeción suelen crecer en forma semicircular, con aspecto de árbol o madera. Pueden ser parásitos, saprótrofos o ambos. Uno de los géneros más comunes, el Ganoderma, puede crecer en grandes y gruesos estantes que pueden contribuir a la muerte del árbol, y luego alimentarse de la madera durante años. Su dureza significa que son muy resistentes y pueden vivir durante mucho tiempo, con muchas especies que incluso desarrollan hermosos círculos multicolores de color que son en realidad anillos de crecimiento anuales. Los políporos se encuentran entre los descomponedores más eficaces de la lignina y la celulosa, los principales componentes de la madera. Debido a esta capacidad, dominan las comunidades de organismos descomponedores de la madera en los ecosistemas terrestres, junto con los hongos corticioides. Al descomponer los troncos de los árboles, reciclan una parte importante de los nutrientes de los bosques.[7]
Sólo se sabe que los basidiomicetos inician la degradación de la lignina (es decir, causan la podredumbre blanca). Un estudio reciente relaciona el fin de la formación de grandes depósitos de carbón a finales del periodo Carbonífero, hace 300 millones de años, con la evolución de los basidiomicetos que degradan la lignina.[8] Una degradación más eficaz de la madera por parte de los hongos significaba que se acumulaba menos material vegetal (y, por tanto, menos carbón) en el suelo.
Por otro lado, la mayoría de los hongos de la podredumbre parda son poliporos. Estas especies han perdido su capacidad de degradación de la lignina pero son muy eficientes en la degradación de la celulosa. Los hongos de podredumbre parda son frecuentes en las coníferas y en los hábitats abiertos y expuestos al sol. La comunidad de hongos en cualquier tronco puede incluir tanto especies de podredumbre blanca como de podredumbre marrón, complementando sus estrategias de degradación de la madera.
Los políporos y otros hongos descomponedores son el primer paso en las cadenas alimentarias que se alimentan de material vegetal descompuesto. Una rica fauna de insectos, ácaros y otros invertebrados se alimenta del micelio de los polporos y de los cuerpos fructíferos, proporcionando además alimento a las aves y otros animales de mayor tamaño. Los pájaros carpinteros y otros pájaros que anidan en agujeros suelen tallar sus nidos en la madera más blanda descompuesta por los políporos.[9]
amenazas
[editar]Casi todos los poliporos dependen de los árboles para su supervivencia. La deforestación y la gestión intensiva de los bosques provocan la disminución de la abundancia y la diversidad de los poliporos. Para muchas especies los cambios pueden ser excesivos, y comienzan un lento deslizamiento hacia la extinción. Dado que la mayoría de las especies de poliporas están relativamente extendidas, este proceso suele ser lento. Las extinciones regionales pueden producirse con relativa rapidez y han sido documentadas (por ejemplo, Antrodia crassa en el norte de Europa).[10]
Los políporos pueden disminuir por muchas razones. Pueden depender de un único huésped o de un hábitat muy especial. Por ejemplo, Echinodontium ballouii sólo se ha encontrado en los pantanos de cedro blanco del Atlántico en el noreste de Estados Unidos.[11][12] Las especies pueden depender de individuos arbóreos muy viejos, como el Bridgeoporus nobilissimus del noroeste de EE UU.[13] Estas dos especies también tienen un área de distribución bastante restringida, lo que las hace más vulnerables a la extinción.
Además del individuo del árbol huésped, las características del hábitat circundante también son importantes. Algunas especies prefieren los bosques de dosel cerrado con un microclima húmedo y homogéneo que podría verse alterado, por ejemplo, por la tala (por ejemplo, Skeletocutis jelicii). Otras sufren la falta de un hábitat forestal abierto en zonas donde se suprimen los incendios (por ejemplo, Gloeophyllum carbonarium en los países nórdicos, donde los incendios forestales forman parte de la dinámica natural del bosque[14]).
Para la mayoría de las especies en declive, el principal problema es la falta de madera muerta en el bosque. Cuando los troncos de los árboles adecuados son demasiado escasos en el paisaje, no todas las especies son capaces de extenderse a nuevos troncos después de que los viejos se hayan consumido, lo que hace que la población disminuya y acabe desapareciendo. Así, las especies que abundan en los bosques antiguos con abundante madera muerta pueden estar totalmente ausentes en los bosques gestionados. Por ejemplo, Amylocystis lapponica y Fomitopsis rosea son especies dominantes en los bosques antiguos de abeto del norte de Europa, desde Polonia hasta Noruega, pero están ausentes en los bosques gestionados.
El cambio climático puede suponer un problema para las poliporas que ya dependen de unos pocos fragmentos de bosques antiguos y pueden ser incapaces de migrar con la vegetación cambiante.
Indicador de valor
[editar]Los políporos se han utilizado como especies indicadoras de bosques naturales sanos o bosques antiguos en Europa. Son buenos indicadores de la diversidad de invertebrados en la madera muerta e incluyen muchas especies en peligro de extinción. Los políporos son buenos indicadores porque son relativamente fáciles de encontrar -muchas especies producen cuerpos fructíferos llamativos y de larga duración- y porque pueden ser identificados en el campo.[15]
La primera lista de indicadores de poliposas ampliamente utilizada en los inventarios forestales y en los trabajos de conservación se elaboró en el norte de Suecia en 1992 (método "Steget före").[16][17] La lista "Steget före" incluía seis políporas en tres clases de valor. En Finlandia, se publicó en 1993 una lista de 30 especies para los bosques dominados por la picea, que fue ampliamente adoptada.[18] Posteriormente se publicó una lista similar para los bosques dominados por pinos. Desde entonces se han publicado en Suecia listas más largas de especies indicadoras.[19][20]
Muchas especies indicadoras están en la lista roja, pero no necesariamente todas. Las listas rojas nacionales de hongos suelen incluir muchos políporos y se utilizan como listas indicadoras del valor de conservación en muchos países europeos.
Clasificación
[editar]Durante la mayor parte del siglo XX, los políporos fueron tratados como una familia, las Polyporaceae. Las reconstrucciones del árbol genealógico de los hongos muestran que el cuerpo fructífero poroide ha evolucionado numerosas veces en el pasado. La clasificación evolutiva moderna basada en el ADN sitúa a los políporos en al menos 12 órdenes.[21][22][23] Los órdenes que contienen la mayoría de las especies de poliporas son los Polyporales (géneros como Fomes, Polyporus y Trametes) y los Hymenochaetales (por ejemplo, Oxyporus, Phellinus y Trichaptum). Desde el punto de vista económico, quizá los poliporos más importantes, Heterobasidion spp., que son plagas de las plantaciones de coníferas, pertenecen a los Russulales.[24] Otros órdenes de poliporas son los Agaricales, Amylocorticiales, Auriculariales, Boletales, Cantharellales, Gloeophyllales, Sebacinales, Thelephorales y Trechisporales.
Los Polyporales, en el sentido moderno, no son sólo los poliporos, sino también otros tipos de cuerpos fructíferos como los hongos de corteza, los hongos hidnoides y los hongos agaricoides. El término polporo que describe un grupo morfológico no debe confundirse con los grupos taxonómicos Polyporales o Polyporaceae de la literatura moderna.
En la actualidad, los políporos se dividen en unos 170 géneros.[25][26] Este número está destinado a aumentar considerablemente gracias a una mejor comprensión de las relaciones evolutivas entre las especies y a la cartografía de la diversidad descubierta en los trópicos. En definitiva, la clasificación de los políporos es cambiante[27]
Delimitación (morfología)
[editar]La mayoría de las poliporas tienen un himenio poroide, pero no todas las especies. Unas pocas, por ejemplo Elmerina holophaea y Lenzites betulina, forman láminas como los agáricos, pero siguen siendo consideradas políporas, ya que en todos los demás aspectos son similares a las políporas estrechamente relacionadas, formando duros cuerpos fructíferos en la madera. Un par de especies en las que los tubos no se han fusionado en forma de panal de abejas se clasifican de forma variable como políporas o no (por ejemplo, Porotheleum fimbriatum). No hay una distinción clara entre los políporos y los hongos hidnoides; algunos políporos con una superficie inferior irregularmente poroide se han considerado tanto políporos como hongos hidnoides (por ejemplo, Echinodontium tinctorium, Irpex lacteus).
Los hongos boleteados son un grupo morfológico separado que no se incluye en los políporos, aunque tengan tubos. Los cuerpos fructíferos carnosos con un tallo y los caracteres microscópicos separan a las boletes de las poliporas.
-
 Irpex lacteus con poros irregulares
Irpex lacteus con poros irregulares -
 Elmerina holophaea, un poliporo con laminas
Elmerina holophaea, un poliporo con laminas


Usos
[editar]Algunas especies de hongos de corchete son comestibles, como el poliesporo de azufre;[28] el hongo lingzhi es otro, que se utiliza en la medicina china. También pueden utilizarse como mecha en una lámpara de aceite/grasa.
El hongo de la yesca (Fomes fomentarius) se ha utilizado como yesca al menos desde la época de Ötzi el Hombre de Hielo. También se ha utilizado para fabricar un material similar al cuero.
El Ganoderma applanatum, la concha del artista, se utiliza como sustrato para los dibujos. Los especímenes frescos desarrollan líneas de color marrón oscuro cuando se dibujan con un estilete. Las líneas se vuelven permanentes cuando el espécimen se seca.[29]
usos medicinales
[editar]La mayoría de los poliporos son comestibles o, al menos, no son tóxicos; sin embargo, un género de poliporos tiene miembros que son venenosos. Los pólporos del género Hapalopilus han provocado envenenamiento en varias personas con efectos que incluyen disfunción renal y desregulación de las funciones del sistema nervioso central.[30] Algunos pólporos se han utilizado en rituales y con fines utilitarios durante mucho tiempo; el famoso Ötzi el Hombre de Hielo fue encontrado portando dos especies diferentes de pólporos: Piptoporus betulinus, notable por su largo uso en la medicina popular europea,[31] y Fomes fomentarius, que probablemente se utilizaba para encender fuego.[32]
Los políporos de hongos medicinales que se utilizan hoy en día son el Ganoderma lucidum coll. (reishi o lingzhi),[33] Trametes versicolor (cola de pavo) y Ganoderma applanatum (Kofuki-saru-no-koshikake japonés). Más allá de su uso tradicional en la fitoterapia, la investigación contemporánea ha sugerido muchas aplicaciones de las poliposas para el tratamiento de enfermedades relacionadas con el sistema inmunitario y la recuperación del cáncer.
Se han estudiado varias especies por su capacidad de producir compuestos con actividad antipatogénica.[34][35]
Véase también
[editar]- Amadou, una sustancia inflamable preparada a partir de hongos de soporte
- Los hongos pleurotoides tienen una forma similar, pero tienen branquias.
- Hongo de la descomposición de la madera
- Lista de los hongos y conks más grandes del mundo
Referencias
[editar]- ↑ «Four Things to Know about Fungi "Climate Warriors"». Boston University (en inglés). Consultado el 27 de noviembre de 2022.
- ↑ «Ainsworth and Bisby's Dictionary of the Fungi (10th edition)». Reference Reviews 23 (5): 42-42. 12 de junio de 2009. ISSN 0950-4125. doi:10.1108/09504120910969104. Consultado el 27 de noviembre de 2022.
- ↑ Phillips, Roger (2006), Mushrooms. Pub. McMilan, ISBN 0-330-44237-6. P. 314.
- ↑ Reid, Derek A. (2006). Mushrooms. Macmillan. ISBN 0-330-44237-6. OCLC 68770488. Consultado el 27 de noviembre de 2022.
- ↑ Alexopoulos, Constantine John; Delevoryas, Theodore (1987). Morphology of plants and fungi (5th ed edición). Harper & Row. ISBN 0-06-040839-1. OCLC 13762074. Consultado el 27 de noviembre de 2022.
- ↑ Gäumann, Ernst Albert; Dodge, Carroll W. (1928). Comparative morphology of Fungi (1st ed. edición). McGraw-Hill Book Company, Inc. Consultado el 27 de noviembre de 2022.
- ↑ Binder, Manfred; Justo, Alfredo; Riley, Robert; Salamov, Asaf; Lopez-Giraldez, Francesc; Sjökvist, Elisabet; Copeland, Alex; Foster, Brian et al. (1 de noviembre de 2013). «Phylogenetic and phylogenomic overview of the Polyporales». Mycologia 105 (6): 1350-1373. ISSN 0027-5514. PMID 23935031. doi:10.3852/13-003. Consultado el 1 de diciembre de 2022.
- ↑ Floudas, Dimitrios; Binder, Manfred; Riley, Robert; Barry, Kerrie; Blanchette, Robert A.; Henrissat, Bernard; Martínez, Angel T.; Otillar, Robert et al. (29 de junio de 2012). «The Paleozoic origin of enzymatic lignin decomposition reconstructed from 31 fungal genomes». Science (New York, N.Y.) 336 (6089): 1715-1719. ISSN 1095-9203. PMID 22745431. doi:10.1126/science.1221748. Consultado el 1 de diciembre de 2022.
- ↑ Kraus, Daniel (2013). Integrative approaches as an opportunity for the conservation of forest biodiversity. European Forest Institute. ISBN 978-952-5980-06-6. OCLC 879248435. Consultado el 1 de diciembre de 2022.
- ↑ Korpilahti, Eeva (2009). «Metsähallituksen 150 vuotta». Metsätieteen aikakauskirja 2009 (2). ISSN 1455-2515. doi:10.14214/ma.5758. Consultado el 1 de diciembre de 2022.
- ↑ Benkert, D. (1987-01). «Gilbertson, R. L. & Ryvarden, L., North American Polypores. Vol. 1 (Abortiporus-Lindtneria). 433 S. 209 Abb. Fungiflora. Oslo, 1986». Feddes Repertorium 98 (1-2): 136-136. ISSN 0014-8962. doi:10.1002/fedr.4910980108. Consultado el 1 de diciembre de 2022.
- ↑ https://web.archive.org/web/20210414043253/http://mushroomhunter.net/ballouii.htm. Archivado desde el original el 14 de abril de 2021. Consultado el 8 de diciembre de 2022. Falta el
|título=(ayuda) - ↑ Horn, Robert B.; Gervais, Jennifer A. (2018-07). «Landscape influence on the local distribution of western pond turtles». Ecosphere 9 (7): e02346. ISSN 2150-8925. doi:10.1002/ecs2.2346. Consultado el 1 de diciembre de 2022.
- ↑ Olsson, Jörgen; Jonsson, Bengt Gunnar (2010-04). «Restoration fire and wood-inhabiting fungi in a Swedish Pinus sylvestris forest». Forest Ecology and Management (en inglés) 259 (10): 1971-1980. doi:10.1016/j.foreco.2010.02.008. Consultado el 1 de diciembre de 2022.
- ↑ Moore, D.; British Mycological Society. Symposium (2001). Fungal conservation : issues and solutions : a special volume of the British Mycological Society. Cambridge University Press. ISBN 978-0-511-56516-8. OCLC 704520157. Consultado el 1 de diciembre de 2022.
- ↑ och Bo Peterson, Gunnel (1972-05). «Svensk Botanisk Tidskrift». Taxon 21 (2/3): 367. ISSN 0040-0262. doi:10.2307/1218228. Consultado el 8 de diciembre de 2022.
- ↑ «Steget Före-metoden». web.archive.org. 11 de agosto de 2010. Archivado desde el original el 11 de agosto de 2010. Consultado el 8 de diciembre de 2022.
- ↑ Suutari, Pekka (1 de mayo de 2003). «Kai Åberg: "Nää laulut kato kertoo meijän elämästä". Tutkimus romanien laulukulttuuristaItä-Suomessa 1990-luvulla. Suomen Etnomusikologisen Seuran julkaisuja 8. Helsinki. 2002.». Elore 10 (1). ISSN 1456-3010. doi:10.30666/elore.78393. Consultado el 8 de diciembre de 2022.
- ↑ «Eksempler på indikatorer i Danmark». dx.doi.org. 4 de marzo de 2014. Consultado el 8 de diciembre de 2022.
- ↑ «Signalarter - Skogsstyrelsen». web.archive.org. 6 de octubre de 2014. Archivado desde el original el 6 de octubre de 2014. Consultado el 8 de diciembre de 2022.
- ↑ Hibbett, David S.; Binder, Manfred; Bischoff, Joseph F.; Blackwell, Meredith; Cannon, Paul F.; Eriksson, Ove E.; Huhndorf, Sabine; James, Timothy et al. (2007-05). «A higher-level phylogenetic classification of the Fungi». Mycological Research 111 (Pt 5): 509-547. ISSN 0953-7562. PMID 17572334. doi:10.1016/j.mycres.2007.03.004. Consultado el 8 de diciembre de 2022.
- ↑ Binder, Manfred; Larsson, Karl-Henrik; Matheny, P. Brandon; Hibbett, David S. (2010-07). «Amylocorticiales ord. nov. and Jaapiales ord. nov.: Early diverging clades of Agaricomycetidae dominated by corticioid forms». Mycologia (en inglés) 102 (4): 865-880. ISSN 0027-5514. doi:10.3852/09-288. Consultado el 8 de diciembre de 2022.
- ↑ Aime, M. Catherine; Henkel, Terry W.; Ryvarden, Leif (2003-07). «Studies in Neotropical Polypores 15: New and Interesting Species from Guyana». Mycologia 95 (4): 614. ISSN 0027-5514. doi:10.2307/3761937. Consultado el 8 de diciembre de 2022.
- ↑ Garbelotto, Matteo; Gonthier, Paolo (4 de agosto de 2013). «Biology, Epidemiology, and Control of Heterobasidion Species Worldwide». Annual Review of Phytopathology 51 (1): 39-59. ISSN 0066-4286. doi:10.1146/annurev-phyto-082712-102225. Consultado el 8 de diciembre de 2022.
- ↑ Benkert, D. (1987-01). «Gilbertson, R. L. & Ryvarden, L., North American Polypores. Vol. 1 (Abortiporus-Lindtneria). 433 S. 209 Abb. Fungiflora. Oslo, 1986». Feddes Repertorium 98 (1-2): 136-136. ISSN 0014-8962. doi:10.1002/fedr.4910980108. Consultado el 8 de diciembre de 2022.
- ↑ «Mycobank». www.mycobank.org. Consultado el 8 de diciembre de 2022.
- ↑ Justo, Alfredo; Miettinen, Otto; Floudas, Dimitrios; Ortiz-Santana, Beatriz; Sjökvist, Elisabet; Lindner, Daniel; Nakasone, Karen; Niemelä, Tuomo et al. (2017-09). «A revised family-level classification of the Polyporales (Basidiomycota)». Fungal Biology (en inglés) 121 (9): 798-824. doi:10.1016/j.funbio.2017.05.010. Consultado el 8 de diciembre de 2022.
- ↑ Kuo, Michael (2007). 100 edible mushrooms. University of Michigan Press. ISBN 978-0-472-03126-9. OCLC 76820806. Consultado el 8 de diciembre de 2022.
- ↑ Evans, Shelley (2011). The book of fungi : a life-size guide to six hundred species from around the world. The University of Chicago Press. ISBN 978-0-226-72117-0. OCLC 664519721. Consultado el 8 de diciembre de 2022.
- ↑ Saviuc, Philippe; Danel, Vincent (2006). «New Syndromes in Mushroom Poisoning:». Toxicological Reviews (en inglés) 25 (3): 199-209. ISSN 1176-2551. doi:10.2165/00139709-200625030-00004. Consultado el 8 de diciembre de 2022.
- ↑ Grienke, Ulrike; Zöll, Margit; Peintner, Ursula; Rollinger, Judith M. (2014-07). «European medicinal polypores – A modern view on traditional uses». Journal of Ethnopharmacology (en inglés) 154 (3): 564-583. doi:10.1016/j.jep.2014.04.030. Consultado el 8 de diciembre de 2022.
- ↑ Peintner, U.; Pöder, R.; Pümpel, T. (1998-10). «The iceman's fungi». Mycological Research (en inglés) 102 (10): 1153-1162. doi:10.1017/S0953756298006546. Consultado el 8 de diciembre de 2022.
- ↑ Bishop, Karen S.; Kao, Chi H.J.; Xu, Yuanye; Glucina, Marcus P.; Paterson, R. Russell M.; Ferguson, Lynnette R. (2015-06). «From 2000years of Ganoderma lucidum to recent developments in nutraceuticals». Phytochemistry (en inglés) 114: 56-65. doi:10.1016/j.phytochem.2015.02.015. Consultado el 8 de diciembre de 2022.
- ↑ Zjawiony, Jordan K. (1 de febrero de 2004). «Biologically Active Compounds from Aphyllophorales (Polypore) Fungi». Journal of Natural Products (en inglés) 67 (2): 300-310. ISSN 0163-3864. doi:10.1021/np030372w. Consultado el 8 de diciembre de 2022.
- ↑ Rai, Mahendra K.; Gaikwad, Swapnil; Nagaonkar, Dipali; dos Santos, Carolina Alves (2015). «Current Advances in the Antimicrobial Potential of Species of Genus Ganoderma (Higher Basidiomycetes) against Human Pathogenic Microorganisms (Review)». International Journal of Medicinal Mushrooms (en inglés) 17 (10): 921-932. ISSN 1521-9437. doi:10.1615/IntJMedMushrooms.v17.i10.20. Consultado el 8 de diciembre de 2022.