Area 10 di Brodmann
| Area di Brodmann 10 | |
|---|---|
 | |
 | |
| Sistema | Sistema nervoso centrale |
| Localizzazione anatomica | cervello |
| Identificatori | |
| FMA | 68607 |
| ID NeuroLex | birnlex_1741 |
L'area 10 di Brodmann, oppure BA10 è la parte frontopolare della corteccia frontale nel cervello umano.[1] BA10 è stata inizialmente definita in termini di tratti di citoarchitettonica nei cervelli autoptici; la ricerca sull'imaging funzionale moderno non può identificare direttamente questi confini ed i termini prefrontale anteriore, corteccia prefrontale rostrale e corteccia prefrontale frontopolare vengono usati per riferirsi all'area nella parte più anteriore della corteccia frontale che si avvicina oppure che copre principalmente la BA10.
L'area di Brodmann 10 è la maggiore area cito-architettonica nel cervello umano. Spesso viene descritta come "una delle regioni del cervello umano meno compresa".[2] La ricerca più attuale suggerisce che sia coinvolta in processi strategici nel recupero della memoria e che serva a coordinare l'ordine di complesse funzioni esecutive. Durante l'evoluzione umana, le funzioni richieste a quest'area, portarono ad un vantaggio dovuto a pressione selettiva degli individui con grosse aree 10, e portarono alla sua graduale espansione nelle dimensioni relative in tutta la popolazione umana, rispetto al minore sovra-sviluppo di altre aree del cervello.[3]
Anatomia
[modifica | modifica wikitesto]Dimensioni
[modifica | modifica wikitesto]Il volume dell'area 10 di Brodmann negli umani è mediamente di 14 cm³ e costituisce circa il 1,2% del volume cerebrale. Questo è il doppio della media di un ominide con un cervello delle dimensioni umane. A confronto, il volume medio dell'area 10 di un bonobo è di 2,8 cm³, ammontando al 0,74% del suo cervello. In ogni emisfero, l'area 10 contiene circa 250 milioni di neuroni.[3]
Localizzazione
[modifica | modifica wikitesto]Quest'area è una suddivisione del lobo frontale della corteccia cerebrale che è stato definito dalla citoarchitettonica. Occupa la porzione più rostrale del giro frontale superiore e del giro frontale medio. Negli esseri umani sulla faccia mediale dell'emisfero si trova circondata ventralmente dal solco rostrale superiore. Non si estende fino ad arrivare al solco cingolato. Citoarchitettonicamente è limitata dorsalmente dalla area 9 di Brodmann, caudalmente dalla area frontale mediale 46, e ventralmente dall'area orbitale 47 e dall'area rostrale 12 oppure, in una versione pristina della mappa corticale di Brodmann (Brodmann-1909), con l'area prefrontale 11-1909.[4]
L'area 10 giace al di dietro del seno frontale paranasale, condizione che pone limiti alla ricerca elettrofisiologica.[5]
Relazione col polo frontale del cranio
[modifica | modifica wikitesto]Negli esseri umani l'area del polo frontale della corteccia prefrontale che include non soltanto l'area 10 ma anche parte dell'area 9 di Brodmann. La BA 10 si estende anche al di là dell'area fronto-polare (elettrodi fP1 e fP2 del sistema 10-20 in elettroencefalografia) verso il suo lato ventro-mediale. Nelle scimmie studiate da Guenon, l'area fronto-polare è riempita dalla BA 12 (e la sua BA 10 si trova nella regione orbitale prefrontale region).[2]
Citoarchitettonica
[modifica | modifica wikitesto]Negli esseri umani i sei strati corticali dell'area 10 sono stati descritti come aventi un "aspetto omogeneo in modo rimarchevole".[3] Anche se tutti sono rapidamente identificabili. Rispetto ad ogni altro, lo strato I è sottile in ampiezza, ammontando all'11% in profondità dell'area 10. Lo strato II è sottile e contiene piccole cellule granulari e piramidali (di dimensioni medie) che si colorano da grigio intermedio a scuro con la tintura di Nissl, che tinge il RNA e il DNA. Lo strato più ampio è quello III. I suoi neuroni piramidali sono più piccoli vicino al sovrastante strato II rispetto al sottostante strato IV. Come lo strato II le sue cellule si colorano da mediamente a scuro. Gli strati II e III ammontano a circa il 43% dello spessore corticale. Lo strato IV, sottile, ha bordi netti che lo separano dagli strati III e V. Con cellule che si colorano da pallido a medio. Lo strato V è ampio, suddivisibile in due sottostrati Va e Vb. La densità di cellule in Va, più scure, è maggiore rispetto a Vb. Gli strati IV e V sono il 40% dello spessore corticale. Lo strato VI, al di sopra delle fibre bianche, contiene cellule piramidali scure e neuroni fusiformi (caratteristici dei grossi cervelli: uomo, balena, delfino), e contribuisce al 6% dello spessore dell'area 10 di Brodmann.[3]
L'area 10 differisce dall'adiacente area 9 di Brodmann perché la seconda ha uno strato Vb che si distingue meglio e uno strato II più prominente. La vicina area 11 di Brodmann ha uno strato IV più sottile con strati Va, Vb e II più spessi.[3]
L'area 10 degli umani ha la più bassa densità neuronale tra i cervelli dei primati.[3] Inusualmente possiede neuroni con un'arborizzazione dendritica particolarmente estesa e altamente densa di spine dendritiche.[6] Alcuni suggeriscono che queste connessioni permettano di integrare informazioni provenienti da molteplici aree.[2]
Subaree
[modifica | modifica wikitesto]L'area 10 di Brodmann viene divisa in tre sotto-aree, 10p, 10m e 10r. La sub-area 10p occupa il polo frontale mentre le altre due coprono la parte ventro-mediale della corteccia prefrontale.[7] L'area 10m ha dei sottili strati II e IV e uno strato V più spesso. All'opposto, l'area 10r ha un prominente strato II e uno strato IV più prominente. Le grosse cellule piramidali sono anche presenti nello strato III della sub-area 10r e anche di più nell'area 10p. Ma si può notare che, "le differenze tra le tre aree sono graduali, comunque, ed è difficile disegnare confini netti tra di esse".[7]
Connessioni
[modifica | modifica wikitesto]La ricerca sui primati suggerisce che l'area 10 ha connessioni in ingresso ed entrata con altre aree associative di alto livello della corteccia, particolarmente nell'ambito della corteccia prefrontale, mentre ne ha ben poche con le aree primarie sensoriali o motorie. Le sue connessioni attraverso la capsula estrema la collegano alle aree uditive e multi-sensoriali del solco temporale superiore. Queste connessioni proseguono verso il fascicolo longitudinale mediale nella sostanza bianca del giro temporale superiore, nel bordo superiore del giro temporale superiore (aree TAa, TS2, e TS3) e nelle vicine aree multi-sensoriali nel bordo superiore del solco temporale superiore (TPO). Un'altra regione collegata alla capsula estrema è la regione ventrale dell'insula. Connessioni attraverso il fascicolo cingolato collegano l'area 10 alla corteccia cingolata anteriore e posteriore e alla corteccia retrospleniale. Il fascicolo uncinato connette con l'amigdala, la neocorteccia temporopolare e gran parte della circonvoluzione temporale superiore anteriore. Non ci sono connessioni con la corteccia parietale, corteccia occipitale e la circonvoluzione temporale inferiore.[8] Non sembra essere connessa con le aree "a valle" come le altre aree prefrontali, è l'unica regione prefrontale che è principalmente (e quasi esclusivamente) connessa con la corteccia supramodale, la corteccia temporale anteriore e la corteccia cingolata.[9] Si pensa che queste connessioni giochino un maggiore ruolo nell'integrazione delle informazioni provenienti dai sistemi visivi, uditivi e somatici in modo tale da ottenere un'interpretazione astratta, amodale e concettuale dell'ambiente. Si pensa che la corteccia prefrontale influenzi l'elaborazione delle informazioni astratte e l'integrazione dei risultati di molteplici operazioni cognitive.[8]
Evoluzione
[modifica | modifica wikitesto]Katerina Semendeferi e i suoi colleghi hanno suggerito che "Durante l'evoluzione degli ominidi, nell'area 10 si sono verificati diversi cambiamenti:si è verificato un considerevole aumento della dimensione complessiva, che ha comportato un incremento della connettività, specialmente con le altre aree d associative di ordine superiore."[3]
Calchi del cranio presi dall'interno del teschio dell’Homo floresiensis mostrano un'espansione della regione polare frontale che suggeriscono un ampliamento dell'area 10 di Brodmann.[10]
Funzioni
[modifica | modifica wikitesto]Anche se questa regione è molto estesa negli esseri umani, la sua funzione è scarsamente compresa. Koechlin & Hyafil hanno proposto che l'elaborazione della ramificazione cognitiva sia la funzione principale della corteccia fronto-polare[11]. La ramificazione cognitiva (o ragionamento relazionale o suddivisione e gerarchia dei compiti) conferisce a un compito che veniva eseguito previamente, uno status di minore importanza e viene messo in attesa, per essere successivamente ricuperato ed eseguito dopo il completamento di quello gerarchicamente superiore. Molti tra i nostri complessi comportamenti e attività mentali richiedono il simultaneo coinvolgimento di molteplici compiti, e suggeriscono che la corteccia prefrontale anteriore possa eseguire una funzione di dominio generale in queste operazioni di elaborazione di liste di compiti successivi e nel determinare loro proprietà e reciproca interdipendenza. Comunque, altre ipotesi sono state formulate, come quelle di Burgess e collaboratori.[12]
Galleria d'immagini
[modifica | modifica wikitesto]-
 Animazione.
Animazione. -
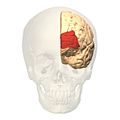 Visione frontale.
Visione frontale. -
 Visione laterale.
Visione laterale. -
 Visione mediale.
Visione mediale.




Note
[modifica | modifica wikitesto]- ^ (EN) Barbara J. Knowlton, Robert G. Morrison e John E. Hummel, A neurocomputational system for relational reasoning, in Trends in Cognitive Sciences, vol. 16, n. 7, 7 gennaio 2012, pp. 373-381, DOI:10.1016/j.tics.2012.06.002. URL consultato il 13 dicembre 2015.
- ^ a b c Ramnani N, Owen AM. (2004). Anterior prefrontal cortex: insights into function from anatomy and neuroimaging. Nat Rev Neurosci. 5(3):184-94. DOI: 10.1038/nrn1343 PMID 14976518
- ^ a b c d e f g Semendeferi K, Armstrong E, Schleicher A, Zilles K, Van Hoesen GW. (2001). Prefrontal cortex in humans and apes: a comparative study of area 10. Am J Phys Anthropol. 114(3):224-41. DOI: 10.1002/ajpa.20947 PMID 11241188
- ^ BrainInfo: area 10 of Brodmann (human).
- ^ Wallis JD. (2010). Polar exploration. Nat Neurosci. 13(1):7-8. DOI: 10.1038/nn0110-7 PMID 20033080
- ^ Jacobs B, Schall M, Prather M, Kapler E, Driscoll L, Baca S, Jacobs J, Ford K, Wainwright M, Treml M. (2001). Regional dendritic and spine variation in human cerebral cortex: a quantitative golgi study.. Cereb Cortex. 11(6):558-71. DOI: 10.1093/cercor/11.6.558 PMID 11375917
- ^ a b Ongür D, Ferry AT, Price JL. (2003). Architectonic subdivision of the human orbital and medial prefrontal cortex. J Comp Neurol. 460(3):425-49. DOI: 10.1002/cne.10609 PMID 12692859
- ^ a b (EN) Michael Petrides e Deepak N. Pandya, Efferent Association Pathways from the Rostral Prefrontal Cortex in the Macaque Monkey, in The Journal of Neuroscience, vol. 27, n. 43, 24 ottobre 2007, pp. 11573-11586, DOI:10.1523/JNEUROSCI.2419-07.2007. URL consultato il 13 dicembre 2015.
- ^ Narender Ramnani e Adrian M. Owen, Anterior prefrontal cortex: insights into function from anatomy and neuroimaging, in Nature Reviews Neuroscience, vol. 5, n. 3, pp. 184-194, DOI:10.1038/nrn1343.
- ^ (EN) Dean Falk, Charles Hildebolt e Kirk Smith, The Brain of LB1, Homo floresiensis, in Science, vol. 308, n. 5719, 8 aprile 2005, pp. 242-245, DOI:10.1126/science.1109727. URL consultato il 13 dicembre 2015.
- ^ Koechlin & Hyafil - Anterior prefrontal function and the limits of human-decision making. Science, Vol. 318, 594-598.
- ^ Burgess, P.W., Dumontheil, I., & Gilbert, S.J. (2007). The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends in Cognitive Science, Vol. 11, No. 7.
Voci correlate
[modifica | modifica wikitesto]Altri progetti
[modifica | modifica wikitesto] Wikimedia Commons contiene immagini o altri file su Area 10 di Brodmann
Wikimedia Commons contiene immagini o altri file su Area 10 di Brodmann
Collegamenti esterni
[modifica | modifica wikitesto]- Contrappunti - Generazione X e la Trappola del Multitasking, su punto-informatico.it.
- genM: The Multitasking Generation, su time.com (archiviato dall'url originale l'11 maggio 2011).
- Brede Database Brodmann area 10, su neuro.imm.dtu.dk. URL consultato il 6 settembre 2010 (archiviato dall'url originale il 19 luglio 2011).
- BrainMaps Area 10 Of Prefrontal Cortex, su brainmaps.org.
- Is Brodmann Area 10 the Key to Human Evolution?, su questioneverything.typepad.com.