Alveolata
| Alveolata (Potcarstvo: Alveolata; Vremenski raspon: Ediakaran; prije ~ 850 miliona godina —→ Sadašnjost | |
|---|---|
| (Vidi IUCN-ov crveni popis za kratice) | |
 | |
| Sistematika | |
| Carstvo | Chromista |
| Potcarstvo | Harosa Infrakoljeno Alveolata Cavalier-Smith, 1991 |
| Myzozoa Ciliophora | |
| Sinonimi | |
| Alveolatobiontes | |
Alveolata (što znači "sa šupljinama)[1] su grupa protista, kao glavnim kladusom[2], natkoljeno[3] unutar Eukarya, također zvanih Alveolata.[4]
Potcarstvo Harosa karakterizira kortikalna alveola, kao spljoštena vezikula ispod ćelijske membrane s kojom čini pelikulu (vanjski „zid“); on im ujedno omogućava stalnost oblika tijela. Postoji podjela na dva koljena: Myzozoa i Cilliata (Ciliophora).[3]
Karakteristike[uredi | uredi izvor]
Najistaknutija zajednička karakteristika je prisustvo kortikalnih (vanregijskih) alveola (vreća). To su spljoštene (vezikulske) vrećice upakovane u kontinuirani sloj tik ispod membrane i podržavaju je, obično formirajući fleksibilnu pelikulu (tanku kožu). U dinoflagelata često formiraju oklopne ploče. Alveolati imaju mitohondrije sa cjevastim kristama (grebenima), a njihove flagele ili cilije imaju izrazitu strukturu. Gotovo svi sekvencionirani mitohondrijski genomi cilijata i apikopleksija su linearni.[5] Mitohondrijsk i genom Babesia microti je kružnog oblika.[6] Sada se zna i da ova vrsta ne pripada nijednom od rodova Babesia ili Theileria i za nju će se morati stvoriti novi rod.
Razvoj[uredi | uredi izvor]
Razvoj plastida među alveolatima je intrigantan. Cavalier-Smith predložio je da su alveolate evoluirale od predaka koji sadrže hloroplaste, što je takođe stvorilo hipotezu Chromistama (hromalveolata). Drugi istraživači nagađaju da su alveolatima prvobitno nedostajali plastidi i da su ih eventualno dinoflagelati i Apicomplexa stekli odvojeno. Međutim, sada se čini da su alveolate, dinoflagelate, Chromerida i heterokontne alge svoje plastide dobili iz crvene alge s dokazima o zajedničkom porijeklu ove organele u sva ova četiri kladusa.[7]
Evolucija[uredi | uredi izvor]
Bayesova procjena postavlja evoluciju alveolatne grupe na od prije ~ 850 miliona godina.[8] Alveolata uključuju Myzozoa, Ciliates i Colponemida. Drugim riječima, termin Myzozoa, što znači "sipati sadržaj iz plijen", može se neformalno primijeniti na zajedničkog pretka podskupina alveolata koji nisu ni cililate ni kolponemide. Predacija algi važan je pokretač u evoluciji alveolata, jer može pružiti izvore za endosimbiozu novih plastida. Termin Myzozoa je stoga koristan za koncept praćenje evolucijske historije koljena Alveolata.
Preci alveolatne grupe možda su bili fotosintetski.[9] Predački alveolati vjerovatno su imali plastide. Hromeridi, apikopleksi i peridininea-dinoflagelati zadržali su ove organele.[10] Napredujući korak dalje, hromeridi, peridininijski dinoflagelati i heterokontne alge imaju zajedničku monofletnu plastidnu liniju, tj. stekli su plastide iz crvenih agi,[7] pa se čini vjerovatnim da je zajednički predak alveolata i heterokonta također bio sposoban za fotosintezu.
U jednoj školi naučnika, zajednički preci dinoflagelata, apikompleksi, Colpodella, Chromerida i Voromona bili su mitocitotske grabljivice s dva heterodinamična biča, mikroporama, trihocistama, mikronemama, polarnim prstenom i zavojnicom koja je otvorena sa strane konoida.[11] Iako je zajednički predak alveolata također mogao imati neke od ovih karakteristika, tvrdi se da mizocitoza nije jedna od tih, jer cilijate gutaju plijen drugačijim mehanizmom.
U toku je rasprava koja se odnosi na broj membrana koje okružuju plastide apikopleksa i određenih dinoflagelata i porijeklo tih membrana. Ovaj ultrastrukturni karakter može se koristiti za grupiranje organizama i ako im je zajednički, to može značiti da su koljena imala zajedničkog fotosintetskog pretka. Na osnovu toga što apikoplekse posjeduju plastid okružen sa četiri membrane, a peridininijski dinoflagelati plastide s tri membrane, Petersen et al.[12] nisu mogli da potvrde da je zajednički stramenopilo-alveolatni plastid mogao biti recikliran više puta u alveolatnom tipu, a izvor su donori stramenopile-alveolata, putem mehanizama gutanja i endosimbioza.
Cilijate su model alveolata, a genetički su detaljno proučavani, tokom najdužeg perioda bilo koje alveolatne loze. Oni su neobični među eukariotima zato što reprodukcija uključuje mikronukleus i makronukleus. Njihova se reprodukcija lahko proučava u laboratorijama, pa ih je historijski učinila uzorima eukariota. Budući da su u potpunosti grabljivice i nedostaju im preostali plastidi, njihova evolucija vrsta ilustrira kako su grabežljivost i autotrofija[9] u dinamičkoj ravnoteži i da ravnoteža može na taj ili na drugi način balansirati na mjestu nastanka novog koljena od miksotrofnih predaka, uzrokujući gubitak jedne sposobnosti.
Klasifikacija[uredi | uredi izvor]
Alveolata se sastoji od oko 9 glavnih i manjih grupa, koje su vrlo raznolike forme, a poznato je da ih povezuju razne ultrastrukturne i genetičke sličnosti:[13]
- Ciliata – vrlo česte protozoe s mnogo kratkih cilija poredanih u redove, te dva jedra
- Acavomonidia[13]
- Colponemidia[13]
- Dinoflagellates s.l. – većinom morske flagelate, od kojih mnoge imaju hloroplaste
- Perkinsozoa
- Chromerida – koljeno morskih fotosintetskih protozoa
- Colpodellida
- Voromonadida
- Apicomplexa – parazitske i sekundarno efotoksintetske protozoe, kojima nedostajuakonemane lokomotivne strukture, osim u gameta
Acavomonidia i Colponemidia su prethodno grupirane kao kolponemidi, takson koji je sada razdeljen na osnovu ultrastrukturne analize. Akavomonidije su bliže grupi dinoflagelata / perkinida nego Colponemidia.
Apicomplexa i dinoflagelati mogu biti bliže jedni drugima nego cilijatima. Obje grupe imaju plastide i većina ih ima snop ili konus od mikrotubula na vrhu ćelije. U apikompleksima to čini dio složene strukture koja se koristi za ulazak u ćelija domaćina, dok u nekim bezbojnim dinoflagelatima formira stabljiku koja se koristi za gutanje plijena. Razni drugi rodovi usko su povezani s ove dvije skupine, uglavnom flagelati sa sličnom apikalnom strukturom. To uključuje članove koji žive slobodno, u rodovima Oxyrrhis i Colponema , te kod parazitske vrste Perkinsus marinus ,[14] Parvilucifera , Rastrimonas i Ellobiopsida s. Direktnom amplifikacijom gena rRNK u morskim pikoplanktonskim uzorcima 2001. godine utvrđeno je prisustvo dvije nove alveolatnae linije, nazvane grupa I i II.[15][16] Grupa I nema kultivirane srodnike, dok je II grupa povezana s dinoflagelatnim parazitom „Amoebophrya“, koji je do sada bio klasificiran u Syndiniales, u dinoflagelatnom redu. Međuodnosi nekih od ovih glavnih skupina sugerisani su tokom 1980-ih, a specifične relacije između sve tri potvrđen je genetskičm studijama, početkom 1990-ih, čemu su ponajviše doprinijeli Gajadhar et al.[17] Cavalier-Smith uveo je formalni naziv Alveolata 1991. godine, iako je u to vrijeme zapravo grupiranje smatrao parafiletskom, a ne a monofiletskom grupom.
Neke studije sugeriraju da haplosporide, uglavnom paraziti morskih beskičmenjaka, mogu pripadati ovdje, ali im nedostaju alveole i sada su smještene u Cercozoa.
Filogenija[uredi | uredi izvor]
Filogenija je rekonstruirana na osnovu kompilacije rezultata više radova.[3][18][19]
| Alveolata |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Nedavna istraživanja pokazuju da su Dinoflagelllata zapravo nastale u Perkinsea, a Perkinsea su se pojavile u Apicomplexa.[20]
Taksonomija[uredi | uredi izvor]
Alveolata Cavalier-Smith 1991 [Alveolatobiontes]
- Koljeno Ciliophora Doflein 1901 stat. n. Copeland 1956 [Ciliata Perty 1852; Infusoria Bütschli 1887; Ciliae, Ciliozoa, Cytoidea, Eozoa, Heterocaryota, Heterokaryota]
- Potkoljeno Postciliodesmatophora Gerassimova & Seravin 1976
- Razred Heterotrichea Stein 1859
- Razred Karyorelictea Corliss 1974
- Potkoljeno Intramacronucleata Lynn 1996
- Razred ?Mesodiniea Chen et al. 2015
- Infrakoljeno Lamellicorticata
- Razred Litostomatea Small & Lynn 1981
- Razred Armophorea Lynn 2004
- Razred Cariacotrichea Orsi et al. 2011
- Razred Spirotrichea Bütschli 1889
- Infrakoljeno Ventrata Cavalier-Smith 2004 [Conthreep Lynn 2012]
- Red ?Discotrichida Chen et al. 2015
- Razred Protocruziea Chen et al. 2015 [Protocruziidia de Puytorac, Grain & Mignot 1987]
- Razred Colpodea Small & Lynn 1981
- Razred Nassophorea Small & Lynn 1981
- Razred Phyllopharyngea de Puytorac et al. 1974
- Razred Prostomatea Schewiakoff 1896
- Razred Plagiopylea Small & Lynn 1985 sensu Lynn 2008
- Razred Oligohymenophorea de Puytorac et al. 1974
- Potkoljeno Postciliodesmatophora Gerassimova & Seravin 1976
- Koljeno Miozoa Cavalier-Smith 1987
- Potkoljeno Colponemidia Tikhonenkov, Mylnikov & Keeling 2013
- Razred Colponemea Cavalier-Smith 1993
- Potkoljeno Acavomonadia Tikhonenkov et al. 2014
- Razred Acavomonadea Tikhonenkov et al. 2014
- Potkoljeno Myzozoa Cavalier-Smith 2004
- Infrakoljeno Apicomplexa Levine 1970 emend. Adl et al. 2005
- Red ?Vitrellida Cavalier-Smith 2017
- Razred ?Myzomonadea Cavalier-Smith & Chao 2004 sensu Ruggiero et al. 2015
- Razred Chromerea
- Razred Colpodellida Patterson & Zölffel 1991 [Spiromonadida Krylov & Mylnikov 1986]
- Nadrazred Sporozoa Leuckart 1879 stat. nov. Cavalier-Smith 2013 [Gamontozoa]
- Razred Blastogregarinida Chatton & Villeneuve 1936 [Blastogregarinina; Blastogregarinorina Chatton & Villeneuve 1936]
- Razred Paragregarea Cavalier-Smith 2014
- Razred Gregarinomorphea Grassé 1953
- Razred Coccidiomorphea Doflein 1901
- Infrakoljeno Dinozoa Cavalier-Smith 1981 emend. 2003
- Red ?Acrocoelida Cavalier-Smith & Chao 2004
- Red ?Rastromonadida Cavalier-Smith & Chao 2004
- Red Squirmidea Norén 1999 stat. nov. Cavalier-Smith 2014
- Nadrazred Perkinsozoa Norén et al. 1999 s.s.
- Razred Perkinsea Levine 1978 [Perkinsasida Levine 1978]
- Nadrazred Dinoflagellata Butschli 1885 stat. nov. Cavalier-Smith 1999 sensu Cavalier-Smith 2013 [Dinozoa Cavalier-Smith 1981]
- Razred Pronoctilucea
- Razred Ellobiopsea Cavalier-Smith 1993 [Ellobiophyceae Loeblich III 1970; Ellobiopsida Whisler 1990]
- Razred Myzodinea Cavalier-Smith 2017
- Razred Oxyrrhea Cavalier-Smith 1987
- Razred Syndinea Chatton 1920 s.l. [Syndiniophyceae Loeblich III 1970 s.s.; Syndina Cavalier-Smith]
- Razred Endodinea Cavalier-Smith 2017
- Razred Noctiluciphyceae Fensome et al. 1993 [Noctilucae Haeckel 1866; Noctilucea Haeckel 1866 stat. nov.; Cystoflagellata Haeckel 1873 stat. nov. Butschli 1887]
- Razred Dinophyceae Pascher 1914 [Peridinea Ehrenberg 1830 stat. nov. Wettstein]
- Infrakoljeno Apicomplexa Levine 1970 emend. Adl et al. 2005
- Potkoljeno Colponemidia Tikhonenkov, Mylnikov & Keeling 2013
Galerija[uredi | uredi izvor]
-

-
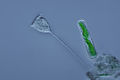 Vorticella (Ciliophora) (lijevo)
Vorticella (Ciliophora) (lijevo) -

-
-






Također pogledajte[uredi | uredi izvor]
Reference[uredi | uredi izvor]
- ^ "alveolate". Memidex (WordNet) Dictionary/Thesaurus. Arhivirano s originala, 11. 4. 2016. Pristupljeno 26. 1. 2011.
- ^ Adl, S.M.; et al. (2012). "The revised classification of eukaryotes". Journal of Eukaryotic Microbiology. 59 (5): 429–514. doi:10.1111/j.1550-7408.2012.00644.x. PMC 3483872. PMID 23020233.
- ^ a b c Ruggiero MA, Gordon DP, Orrell TM, Bailly N, Bourgoin T, Brusca RC, Cavalier-Smith T, Guiry MD, Kirk PM (2015). "A higher level classification of all living organisms". PLoS ONE. 10 (4): e0119248. doi:10.1371/journal.pone.0119248. PMC 4418965. PMID 25923521.CS1 održavanje: upotreba parametra authors (link)
- ^ Cavalier-Smith, T. (1991). "Cell diversification in heterotrophic flagellates". u Patterson, David J.; Larsen, Jacob; Systematics Association (ured.). The Biology of free-living heterotrophic flagellates. Oxford University Press. str. 113–131. ISBN 978-0-19-857747-8.
- ^ Barth, D; Berendonk, TU (2011). "The mitochondrial genome sequence of the ciliate Paramecium caudatum reveals a shift in nucleotide composition and codon usage within the genus Paramecium". BMC Genomics. 12: 272. doi:10.1186/1471-2164-12-272. PMC 3118789. PMID 21627782.
- ^ Cornillot E, Hadj-Kaddour K, Dassouli A, Noel B, Ranwez V, Vacherie B, Augagneur Y, Brès V, Duclos A, Randazzo S, Carcy B, Debierre-Grockiego F, Delbecq S, Moubri-Ménage K, Shams-Eldin H, Usmani-Brown S, Bringaud F, Wincker P, Vivarès CP, Schwarz RT, Schetters TP, Krause PJ, Gorenflot A, Berry V, Barbe V, Ben Mamoun C (2012). "Sequencing of the smallest Apicomplexan genome from the human pathogen Babesia microti". Nucleic Acids Res. 40 (18): 9102–14. doi:10.1093/nar/gks700. PMC 3467087. PMID 22833609.CS1 održavanje: upotreba parametra authors (link)
- ^ a b Janouskovec, J; Horák, A; Oborník, M; Lukes, J; Keeling, PJ (2010). "A common red algal origin of the apicomplexan, dinoflagellate, and heterokont plastids". Proc Natl Acad Sci USA. 107 (24): 10949–54. doi:10.1073/pnas.1003335107. PMC 2890776. PMID 20534454.
- ^ Berney, C; Pawlowski, J (2006). "A molecular time-scale for eukaryote evolution recalibrated with the continuous microfossil record". Proc Biol Sci. 273 (1596): 1867–72. doi:10.1098/rspb.2006.3537. PMC 1634798. PMID 16822745.
- ^ a b Reyes-Prieto, A; Moustafa, A; Bhattacharya, D (2008). "Multiple genes of apparent algal origin suggest ciliates may once have been photosynthetic". Curr. Biol. 18 (13): 956–62. doi:10.1016/j.cub.2008.05.042. PMC 2577054. PMID 18595706.
- ^ Moore RB, Oborník M, Janouskovec J, Chrudimský T, Vancová M, Green DH, Wright SW, Davies NW, Bolch CJ, Heimann K, Slapeta J, Hoegh-Guldberg O, Logsdon JM, Carter DA (2008). "A photosynthetic alveolate closely related to apicomplexan parasites". Nature. 451 (7181): 959–963. doi:10.1038/nature06635. PMID 18288187.CS1 održavanje: upotreba parametra authors (link)
- ^ Kuvardina, ON; Leander, BS; Aleshin, VV; Myl'nikov, AP; Keeling, PJ; Simdyanov, TG (2002). "The phylogeny of colpodellids (Alveolata) using small subunit rRNA gene sequences suggests they are the free living sister group to apicomplexans". J Eukaryot Microbiol. 49 (6): 498–504. doi:10.1111/j.1550-7408.2002.tb00235.x. PMID 12503687. Referenca sadrži prazan nepoznati parametar:
|1=(pomoć) - ^ Petersen J, Ludewig AK, Michael V, Bunk B, Jarek M, Baurain D, Brinkmann H (2014). "Chromera velia, endosymbioses and the rhodoplex hypothesis—plastid evolution in cryptophytes, alveolates, stramenopiles, and haptophytes (CASH lineages)". Genome Biol Evol. 6 (3): 666–684. doi:10.1093/gbe/evu043. PMC 3971594. PMID 24572015.
- ^ a b c Tikhonenkov, DV; Janouškovec, J; Mylnikov, AP; Mikhailov, KV; Simdyanov, TG; Aleoshin, VV; Keeling, PJ (2014). "Description of Colponema vietnamica sp.n. and Acavomonas peruviana n. gen. n. sp., two new alveolate phyla (Colponemidia nom. nov. and Acavomonidia nom. nov.) and their contributions to reconstructing the ancestral state of alveolates and eukaryotes". PLoS ONE. 9 (4): e95467. doi:10.1371/journal.pone.0095467. PMC 3989336. PMID 24740116.
- ^ Zhang, H; Campbell, DA; Sturm, NR; Dungan, CF; Lin, S (2011). "Spliced leader RNAs, mitochondrial gene frameshifts and multi-protein phylogeny expand support for the genus Perkinsus as a unique group of Alveolates". PLOS ONE. 6 (5): e19933. doi:10.1371/journal.pone.0019933. PMC 3101222. PMID 21629701.
- ^ López-García P, Rodríguez-Valera F, Pedrós-Alió C, Moreira D (2001). "Unexpected diversity of small eukaryotes in deep-sea Antarctic plankton". Nature. 409 (6820): 603–7. doi:10.1038/35054537. PMID 11214316.
- ^ Moon-van der Staay SY, De Wachter R, Vaulot D (2001). "Oceanic 18S rDNA sequences from picoplankton reveal unsuspected eukaryotic diversity". Nature. 409 (6820): 607–10. doi:10.1038/35054541. PMID 11214317.
- ^ Gajadhar, A. A.; et al. (1991). "Ribosomal RNA sequences of Sarcocystis muris, Theilera annulata, and Crypthecodinium cohnii reveal evolutionary relationships among apicomplexans, dinoflagellates, and ciliates". Molecular and Biochemical Parasitology. 45 (1): 147–153. doi:10.1016/0166-6851(91)90036-6. PMID 1904987.
- ^ Silar, Philippe (2016), "Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes", HAL Archives-ouvertes: 1–462
- ^ Cavalier-Smith, Thomas (5. 9. 2017). "Kingdom Chromista and its eight phyla: a new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences". Protoplasma. 255 (1): 297–357. doi:10.1007/s00709-017-1147-3. PMC 5756292. PMID 28875267.
- ^ Strassert, Jürgen F H; Karnkowska, Anna; Hehenberger, Elisabeth; Campo, Javier del; Kolisko, Martin; Okamoto, Noriko; Burki, Fabien; Janouškovec, Jan; Poirier, Camille (2018) [published online 10 October 2017]. "Single cell genomics of uncultured marine alveolates shows paraphyly of basal dinoflagellates". The ISME Journal (jezik: engleski). 12 (1): 304–308. doi:10.1038/ismej.2017.167. ISSN 1751-7370. PMC 5739020. PMID 28994824.
Vanjski linkovi[uredi | uredi izvor]
| Commons ima datoteke na temu: Alveolata |
- Tree of Life: Alveolates Arhivirano 28. 11. 2020. na Wayback Machine