Lịch sử sự sống
| Một phần của loạt bài viết về |
| Tiến hóa sinh học |
|---|
 |
Lịch sử tiến hóa của sự sống trên Trái Đất là quá trình mà cả sinh vật sống lẫn sinh vật hóa thạch tiến hóa kể từ khi sự sống xuất hiện trên hành tinh, cho tới hiện tại. Trái Đất hình thành khoảng 4,5 tỉ năm trước và các bằng chứng cho thấy sự sống hình thành trước 3,7 tỉ năm.[1][2][3] Mặc dù có một số bằng chứng gợi ra rằng sự sống xuất hiện sớm tận 4,1 tới 4,28 tỉ năm trước, bằng chứng này vẫn gây tranh cãi do các cơ chế phi sinh học có thể đã tạo nên những hóa thạch này.[1][4][5][6][7] Sự tương đồng giữa tất cả các sinh vật ngày nay ám chỉ rằng sự tồn tại của một tổ tiên chung mà tất cả các sinh vật đã biết đều tách ra từ nó thông qua quá trình tiến hóa.[8] Ước tính hơn 99 phần trăm tất cả các loài, lên đến hơn năm tỉ loài,[9] từng sinh sống trên Trái Đất đã tuyệt chủng.[10][11] Các ước tính về số lượng các loài hiện tại của Trái Đất dao động từ 10 triệu tới 14 triệu,[12][13] trong đó ước tính 1,9 triệu đã được định danh[14] và 1,6 triệu được ghi nhận trong cơ sở dữ liệu trung tâm cho tới hiện tại.[15] Gần đây hơn, vào tháng 5 năm 2016, các nhà khoa học đã báo rằng 1 triệu triệu loài được ước tính sống trên Trái Đất vào thời điểm hiện tại với chỉ một phần một nghìn của một phần trăm được mô tả.[16]
−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||
Bằng chứng sớm nhất về sự sống trên Trái Đất là các dấu hiệu cacbon có nguồn gốc sinh vật[3] và hóa thạch stromatolit[17] được phát hiện trong những tảng đá sau trầm tích 3,7 tỉ năm tuổi được tìm thấy ở phía tây Greenland. Vào năm 2015, "tàn tích của đời sống sinh vật" được phát hiện ra có khả năng nằm trong những tảng đá 4,1 tỉ năm tuổi tại Tây Úc.[18][19] Vào tháng 3 năm 2017, các nhà nghiên cứu đã báo cáo về bằng chứng của thứ có khả năng là dạng sống cổ nhất trên Trái Đất. Thứ được cho là các vi sinh vật hóa thạch đã được phát hiện ra trong kết tủa miệng phun thủy nhiệt tại Vành đai Greenstone của Nuvvuagittuq thuộc Quebec, Canada, chúng có lẽ đã sống sớm nhất là 4,280 tỉ năm trước, và không lâu sau sự hình thành của Trái Đất 4,54 tỉ năm trước.[20][21][22] Theo nhà sinh vật học Stephen Blair Hedges, "Nếu sự sống xuất hiện tương đối nhanh trên Trái Đất... thì điều đó có thể phổ biến trong vũ trụ."[18]
Thảm vi sinh vật gồm vi khuẩn và cổ khuẩn cùng sinh sống là dạng sống thống trị vào đầu Liên đại Thái cổ và nhiều bước quan trọng trong quá trình tiến hóa thời kì đầu được các nhà khoa học nghĩ rằng đã diễn ra bên trong chúng.[23] Sự tiến hóa của quang hợp, khoảng 3,5 tỉ năm trước, cuối cùng đã dẫn tới một sự tích tụ sản phẩm thải của nó, khí oxy, trong khí quyển, dẫn tới thảm họa oxy, bắt đầu khoảng 2,4 tỉ năm trước.[24] Bằng chứng sớm nhất của sinh vật nhân thực (tế bào phức tạp với bào quan) có niên đại từ 1,85 tỉ năm trước,[25][26] và trong khi chúng có thể đã có mặt từ sớm hơn như thế, sự đa dạng hóa của chúng đã tăng tốc khi chúng bắt đầu sử dụng khí oxy trong quá trình trao đổi chất của mình. Sau đó, khoảng 1,7 tỉ năm trước, sinh vật đa bào bắt đầu xuất hiện, với các tế bào phân hóa thực hiện các chức năng chuyên biệt.[27] Sinh sản hữu tính, quá trình yêu cầu sự kết hợp của tế bào sinh sản đực và cái (giao tử) để tạo ra một hợp tử trong một quá trình gọi là thụ tinh là, đối nghịch với sinh sản vô tính, phương pháp sinh sản chính cho đại đa số các sinh vật có thể nhìn bằng mắt thường, bao gồm hầu hết tất cả sinh vật nhân thực (tức bao gồm cả động vật và thực vật).[28] Tuy nhiên nguồn gốc và sự tiến hóa của sinh sản hữu tính vẫn còn là một câu đố đối với các nhà sinh học mặc dù nó đúng là đã tiến hóa từ một tổ tiên chung là một loài nhân thực đơn bào.[29] Động vật đối xứng hai bên, loại động vật có mặt trước và mặt sau, xuất hiện vào khoảng 555 triệu năm trước.[30]
Thực vật trên cạn phức tạp sớm nhất có niên đại vào khoảng 850 triệu năm trước,[31] từ các đồng vị cacbon trong đá thuộc kỉ Tiền Camri, trong khi các loài thực vật trên cạn đa bào dạng tảo thì có niên đại thậm chí là vào khoảng 1 tỉ năm trước,[32] mặc dù bằng chứng đã gợi ra rằng các vi sinh vật đã tạo nên các hệ sinh thái trên cạn sớm nhất, ít nhất 2,7 tỉ năm trước.[33] Các vi sinh vật được cho là đã lát đường cho sự khởi đầu của thực vật trên cạn vào Kỷ Ordo. Thực vật trên cạn quá thành công tới nỗi chúng được cho là đã góp phần vào sự kiện tuyệt chủng Devon muộn.[34] (Chuỗi quan hệ nhân quả dài có vẻ như đã bao gồm cả sự thành công của loài cây cổ đại archaeopteris (1) kéo mức CO2 xuống, dẫn tới hiện tượng mát dần toàn cầu và giảm mực nước biển, (2) rễ cây archaeopteris đã nuôi dưỡng sự phát triển của đất, điều này đã làm tăng hiện tượng phong hóa đá, và việc thoát chất dinh dưỡng vào đất xảy ra sau đó có thể đã gây ra hiện tượng nước nở hoa dẫn tới những sự kiện thiếu oxy khiến sự sống dưới đại dương bị chết đi. Các loài dưới biển là nạn nhân chính của sự kiện tuyệt chủng Devon muộn.)
Khu hệ Ediacara xuất hiện trong thời kỳ Kỷ Ediacara,[35] trong khi động vật có xương sống, cùng với hầu hết các ngành hiện đại khác có nguồn gốc vào khoảng 525 Ma trong cuộc bùng nổ kỷ Cambri.[36] Trong suốt Kỷ Permi, động vật Một cung bên, bao gồm tổ tiên của các loài động vật có vú, đã thống trị mặt đất,[37] nhưng hầu hết những thành viên của nhóm này đã tuyệt chủng trong sự kiện tuyệt chủng kỷ Permi-kỷ Trias 252 Ma.[38] Trong quá trình hồi phục từ thảm họa này, thằn lằn chúa trở thành động vật không xương sống trên cạn dồi dào nhất;[39] một nhóm thằn lằn chúa, khủng long, đã thống trị thời kỳ Kỷ Jura và Creta.[40] Sau khi sự kiện tuyệt chủng kỷ Phấn Trắng - Cổ Cận 66 Ma giết đi những loài khủng long không biết bay,[41] động vật có vú nhanh chóng tăng trưởng về kích cỡ và sự đa dạng.[42] Những sự kiện tuyệt chủng hàng loạt như thế có thể đã thúc đẩy quá trình tiến hóa bằng việc cung cấp cơ hội đa dạng hóa cho những nhóm sinh vật mới.[43]
Lịch sử ban đầu của Trái Đất
[sửa | sửa mã nguồn]Lịch sử của Trái Đất và sự sống trên Trái Đất | ||||||||||||||||||||||||
−4500 — – −4000 — – −3500 — – −3000 — – −2500 — – −2000 — – −1500 — – −1000 — – −500 — – 0 — |
| |||||||||||||||||||||||
Scale: Ma (Triệu năm) | ||||||||||||||||||||||||
Những mảnh vẫn thạch cổ nhất tìm thấy trên Trái Đất có tuổi vào khoảng 4,54 tỉ năm; điều này, gắn liền chủ yếu với lớn trầm tích chì cổ, đã giúp ước lượng tuổi Trái Đất vào quanh thời điểm ấy.[46] Mặt trăng có cùng thành phần với lớp vỏ của Trái Đất nhưng lại không có lõi giàu sắt giống như Trái Đất. Nhiều nhà khoa học nghĩ rằng khoảng 40 triệu năm sau khi Trái Đất hình thành, nó đã va chạm với một thiên thể có kích cỡ Sao Hỏa, ném những nguyên liệu vụn vào quỹ đạo và hình thành nên Mặt trăng. Một giả thuyết khác là Trái Đất và Mặt trăng bắt đầu kết tụ lại cùng một lúc nhưng vì Trái Đất có lực hấp dẫn mạnh hơn Mặt trăng thuở ban đầu nên đã hút gần như toàn bộ các hạt sắt trong khu vực.[47]
Cho tới năm 2001, những viên đá cổ nhất được tìm thấy trên Trái Đất có tuổi đời khoảng 3,8 tỉ năm,[48][49][50][51] dẫn các nhà khoa học tới ước tính rằng cho tới thời điểm đó bề mặt Trái Đất đã nằm trong tình trạng nóng chảy. Do đó, họ đặt tên phần lịch sử của Trái Đất này là Liên đại Hỏa thành.[52] Tuy nhiên, việc phân tích các zircon hình thành 4,4 tỉ năm trước đã ám chỉ rằng lớp vỏ Trái Đất đông cứng khoảng 100 triệu năm sau khi hành tinh hình thành và rằng hành tinh nhanh chóng có đại dương và một bầu khí quyển, thứ có thể đã có khả năng hỗ trợ sự sống.[53][54][55]
Bằng chứng từ Mặt Trăng chỉ ra rằng từ 4 tới 3,8 tỉ năm trước nó đã trải qua một đợt đánh bom nặng nề muộn (Late Heavy Bombardment) bởi các mảnh vụn còn sót lại sau sự hình thành của Hệ Mặt Trời, và Trái Đất đáng lẽ đã trải qua một vụ đánh bom thậm chí còn nặng nề hơn do có lực hấp dẫn lớn hơn.[56] Trong khi không có một bằng chứng trực tiếp nào về các điều kiện trên Trái Đất 4 tới 3,8 tỉ năm trước, chẳng có lý do nào để nghĩ rằng Trái Đất cũng bị ảnh hưởng bởi vụ đánh bom nặng nề muộn này.[57] Sự kiện này có lẽ đã lột sạch sẽ bất kỳ bầu khí quyển hay đại dương nào tồn tại trước đó; trong trường hợp này các chất khí và nước từ các vụ va chạm sao chổi có thể đã góp phần vào sự thay thế của chúng, mặc dù khí thải ra từ núi lửa trên Trái Đất đã cung cấp ít nhất một nửa.[58] Tuy nhiên, vào thời điểm này nếu đời sống vi trùng dưới bề mặt đã tiến hóa thì nó sẽ sống sót qua được vụ đánh bom.[59]
Bằng chứng sớm nhất về sự sống trên Trái Đất
[sửa | sửa mã nguồn]Những sinh vật xác định được sớm nhất thì rất nhỏ và tương đối không có nét gì đặc biệt, và hóa thạch của chúng trông giống những cái que nhỏ, do đó rất khó để phân biệt các cấu trúc nổi lên thông qua các quá trình vật lý vô sinh. Bằng chứng không thể tranh cãi cổ nhất về sự sống trên Trái Đất, được diễn giải là vi khuẩn hóa thạch, thì có niên đại 3 tỉ năm.[60] Các phát hiện khác trong đá có niên đại khoảng 3,5 tỉ năm trước đã được diễn giải là vi khuẩn,[61] với bằng chứng địa hóa học cũng có vẻ như cho thấy sự hiện diện của sự sống 3,8 tỉ năm trước.[62] Tuy nhiên, những phân tích này đã được nghiên cứu kĩ lưỡng cẩn thận, và các quá trình không sinh học được tìm thấy có khả năng đã sản sinh ra tất cả các "dấu hiệu của sự sống" từng được báo cáo.[63][64] Trong khi điều này không chứng minh rằng các cấu trúc tìm thấy được có nguồn gốc không sinh học, không thể coi chúng là bằng chứng rõ ràng cho sự tồn tại của sự sống. Các dấu hiệu địa hóa học từ đá lắng đọng 3,4 tỉ năm trước đã được diễn giải là bằng chứng của sự sống,[60][65] mặc dù những phát biểu này vẫn không được xem xét toàn diện bởi các nhà phê bình.
Bằng chứng về các vi sinh vật hóa thạch được coi là có tuổi đời 3,770 triệu tới 4,280 triệu năm được tìm thấy ở Nuvvuagittuq Greenstone Belt tại Quebec, Canada,[20] mặc dù bằng chứng này bị tranh cãi là không thuyết phục.[66]
Nguồn gốc của sự sống trên Trái Đất
[sửa | sửa mã nguồn]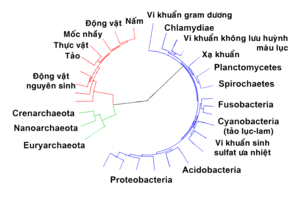
Các nhà sinh vật học lý luận rằng tất cả các sinh vật sống trên Trái Đất hẳn phải có cùng một tổ tiên chung sau cùng duy nhất, bởi vì hầu như không thể có chuyện hai hoặc nhiều nòi giống tách biệt có thể phát triển nhiều những cơ chế sinh hóa phức tạp phổ biến trong tất cả các loài sinh vật sống một cách độc lập được.[68][69]
Xuất hiện độc lập trên Trái Đất
[sửa | sửa mã nguồn]Sự sống trên Trái Đất thì dựa vào cacbon và nước. Cacbon cung cấp một cái khung vững chắc cho các chất hóa học phức tạp và có thể dễ dàng lấy nó ra từ môi trường, đặc biệt là từ cacbon dioxide.[55] Không có nguyên tố hóa học nào khác có thuộc tính đủ giống với cacbon để được coi là thứ tương tự; silic, nguyên tố ở ngay bên dưới cacbon trong bảng tuần hoàn, không tạo thành rất nhiều các nguyên tử bền phức tạp, và bởi vì hầu hết hợp chất của nó không tan trong nước, các sinh vật sẽ khó triết xuất nó ra hơn. Các nguyên tố bo và phosphor có hóa học phức tạp hơn nhưng lại chịu những giới hạn khác khi so với cacbon. Nước là một dung môi xuất sắc và có hai thuộc tính hữu dụng khác: việc băng nổi trên mặt nước giúp cho các sinh vật biển tồn tại bên dưới nó trong mùa đông; và các nguyên tử của nó có cả cực điện tích âm và dương, cho phép nó hình thành hợp chất một cách đa dạng hơn so với các loại dung môi khác. Các loại dung môi tốt khác, ví dụ như ammoniac, chỉ tồn tại dưới dạng lỏng khi ở nhiệt độ thấp tới nỗi mà phản ứng hóa học có lẽ là quá chậm để có thể duy trì sự sống, và thiếu những lợi thế khác của nước.[70] Tuy nhiên, các sinh vật sống dựa vào hóa sinh thay thế có thể có khả năng tồn tại trên các hành tinh khác.[71]
Nghiên cứu về cách mà sự sống có khả năng đã nổi lên từ các chất hóa học vô sinh tập trung vào ba điểm bắt đầu có khả năng xảy ra: tự sao chép, theo đó sinh vật có khả năng sản sinh ra con cháu mà cực kỳ giống với chính nó; trao đổi chất, khả năng ăn và tự sửa chữa chính nó; và màng tế bào bên ngoài, thứ cho phép thức ăn vào trong vào chất thải đi ra ngoài, nhưng không bao gồm những vật chất không mong muốn.[72] Nghiên cứu về hiện tượng tự sinh thì vẫn còn nhiều việc phải làm, vì các hướng tiếp cận theo lý thuyết và theo thực nghiệm thì chỉ mới bắt đầu tiếp xúc với nhau.[70][73]
Sao chép trước tiên: thế giới RNA
[sửa | sửa mã nguồn]Kể cả những thành viên đơn giản nhất của ba vực sự sống hiện đại cũng sử dụng DNA để ghi lại "công thức" của chúng và một dãy phức tạp gồm RNA và các phân tử protein để "đọc" những chỉ dẫn này và sử dụng chúng để phát triển, duy trì và tự sao chép. Việc phát hiện ra rằng một số nguyên tử RNA có thể xúc tác cho cả quá trình sao chép của chính nó lẫn việc tạo ra protein đã dẫn tới giả thuyết về một dạng sống ban đầu dựa hoàn toàn trên RNA.[74] Những ribozyme này có thể đã hình thành nên một thế giới RNA mà trong đó có những cá thể nhưng không có loài, vì đột biến và chuyển gien theo chiều ngang sẽ có nghĩa là con cái ở mỗi thế hệ có khả năng cao sẽ có bộ gien khác với của bố mẹ chúng.[75] RNA sau đó sẽ bị thay thế bởi DNA, thứ ổn định hơn và do đó có thể xây dựng bộ gien dài hơn, mở rộng những tiềm năng mà một sinh vật đơn lẻ có thể có.[76][77] Ribozyme vẫn là thành phần chính của ribosome, "nhà máy protein" của tế bào hiện đại.[78] Bằng chứng gợi ra rằng phân tử RNA đầu tiên hình thành trên Trái Đất vào 4,17 tỉ năm trước.[79]
Mặc dù các phân tử RNA tự sao chép ngắn đã được sản xuất nhân tạo trong phòng thí nghiệm,[76] đã có những nghi ngờ cho rằng ở đâu có thể có quá trình tổng hợp RNA không sinh học tự nhiên.[77] "Ribozyme" sớm nhất có thể đã được hình thành dưới dạng những axit nucleic đơn giản hơn như APN, ATN hoặc AGN, những loại sau đó được thay thế bởi RNA.[78][80]
Vào năm 2003, đã có đề xuất cho rằng kết tủa sulfide kim loại xốp sẽ trợ giúp quá trình tổng hợp RNA ở khoảng 100 °C (212 °F) và áp suất đáy đại dương gần các miệng phun thủy nhiệt. Dưới giả thuyết này, màng lipid sẽ là thành phần tế bào chính xuất hiện cuối cùng và, cho tới lúc đó, các tiền tế bào sẽ bị giam trong các lỗ hổng.[81]
Trao đổi chất trước tiên: thế giới sắt-sulfur
[sửa | sửa mã nguồn]Một chuỗi các thí nghiệm bắt đầu vào năm 1997 đã cho thấy rằng có thể đạt được các giai đoạn đầu của sự hình thành protein từ các vật chất không hữu cơ bao gồm carbon monoxit và hydrogen sulfide bằng cách sử dụng iron sulfide và nickel sulfide làm chất xúc tác. Hầu hết các bước yêu cầu nhiệt độ vào khoảng 100 °C (212 °F) và áp suất vừa phải, mặc dù một giai đoạn cần nhiệt độ 250 °C (482 °F) và áp suất tương đương với áp suất tìm thấy dưới 7 kilômét (4,3 mi) đá. Do đó nó gợi ra rằng việc tổng hợp protein tự duy trì có thể đã xảy ra gần các miệng phun thủy nhiệt.[82]
Màng trước tiên: Thế giới lipid
[sửa | sửa mã nguồn]
Đã có đề xuất là "quả bóng" lipid hai lớp giống như cái mà hình thành nên lớp màng bên ngoài của tế bào có lẽ đã là bước cốt yếu đầu tiên.[83] Các thí nghiệm mô phỏng điều kiện của Trái Đất thời kì đầu đã báo cáo thấy sự hình thành của lipid, và chúng có thể tự động hình thành nên liposome, những "quả bóng" có hai lớp bao bọc, và rồi tự tái tạo.[55] Mặc dù về bản chất chúng không phải là thứ mang thông tin như axit nucleic, chúng sẽ phải phụ thuộc vào chọn lọc tự nhiên cho sự trường thọ và sinh sản. Các loại axit nucleic ví dụ như RNA có lẽ sau đó đã được hình thành một cách dễ dàng hơn bên trong liposome so với việc hình thành ở bên ngoài.[84]
Giả thuyết đất sét
[sửa | sửa mã nguồn]RNA thì phức tạp và có hoài nghi về việc liệu nó có thể xuất hiện theo cách phi sinh học trong tự nhiên.[77] Một số loại đất sét, đáng chú ý là montmorillonit, có các thuộc tính khiến chúng trở thành chất gia tốc đáng tin cậy cho sự nổi lên của một thế giới RNA: chúng phát triển bằng cách tự nhân đôi kiểu mô hình tinh thể của chúng; chúng tuân theo một cái tương tự như chọn lọc tự nhiên, trong đó "loài" đất sét phát triển nhanh nhất trong một môi trường cụ thể thì nhanh chóng trở nên vượt trội; và chúng có thể xúc tác cho quá trình hình thành của các phân tử RNA.[85] Mặc dù ý tưởng này chưa trở thành đồng thuận khoa học, nó vẫn có các nhà ủng hộ tích cực.[86]
Nghiên cứu trong năm 2003 báo cáo rằng montmorillonite cũng có thể đẩy nhanh quá trình biến đổi axit béo thành "bong bóng", và rằng những "bong bóng" đó có thể đóng gói những RNA được đính vào đất sét. Những "bong bóng" này sau đó có thể phát triển bằng cách hấp thụ lipid được bổ sung và rồi sau đó phân chia. Sự hình thành của các tế bào sớm nhất có thể đã được trợ giúp bởi những quá trình tương tự như vậy.[87]
Một giả thuyết tương tự giới thiệu đất sét giàu sắt tự nhân đôi với tư cách là tổ tiên của nucleotide, lipid và amino acid.[88]
Sự sống "nảy mầm" từ nơi nào đó khác
[sửa | sửa mã nguồn]Thuyết tha sinh không giải thích cái cách mà sự sống nảy sinh ngay từ đầu mà chỉ đơn giản là xem xét khả năng nó tới từ một nơi nào đó khác mà không phải Trái Ðất. Cái ý tưởng rằng sự sống trên Trái Ðất được "nảy mầm" từ nơi nào đó khác trong vũ trụ thì đã tồn tại ít nhất từ thời nhà triết học người Hy Lạp Anaximander vào thế kỷ thứ VI SCN.[89] Trong thế kỷ XX đã có đề xuất bởi nhà hóa học vật lý Svante Arrhenius,[90] bởi nhà thiên văn học Fred Hoyle và Chandra Wickramasinghe,[91] và bởi nhà sinh học phân tử Francis Crick và nhà hóa học Leslie Orgel.[92]
Có ba phiên bản chính của giả thuyết "nảy mầm từ nơi khác": từ một nơi khác trong Hệ Mặt Trời của chúng ta thông qua các mảnh vỡ bị bắn vào không gian bởi một vụ va chạm sao chổi lớn, trong tình huống đó thì nguồn gốc đáng tin cậy nhất là Sao Hỏa[93] và Sao Kim;[94] bởi các vị khách người ngoài hành tinh, có thể là kết quả của việc các vi sinh vật lây nhiễm liên hành tinh được chúng mang tới;[92] và từ bên ngoài Hệ Mặt Trời nhưng bằng các phương thức tự nhiên.[90][93]
Các thí nghiệm ở quỹ đạo Trái Đất thấp, ví dụ như EXOSTACK, mô tả rằng một số mầm mống vi sinh vật có thể đã sống sót qua cú sốc của việc bị phóng vào không gian và một số có thể đã sống sót khi bị phơi nhiễm với phóng xạ ngoài không gian trong ít nhất 5,7 năm.[95][96] Các nhà khoa học thì bị giằng co giữa khả năng sự sống phát triển độc lập trên Sao Hỏa,[97] với việc ở trên hành tinh khác trong thiên hà của chúng ta.[93]
Tác động của các cộng đồng vi sinh vật đến tiến hóa và môi trường
[sửa | sửa mã nguồn]
Thảm vi sinh vật (Microbial mat) là các khuẩn lạc đa lớp, đa loài vi khuẩn và các sinh vật khác thường chỉ dày tới độ vài mm, nhưng chứa nhiều loại môi trường hóa học, mỗi một môi trường này phù hợp với bộ các loài vi sinh khác nhau.[98] Ở một mức độ nào đó, mỗi thảm vi sinh tạo thành chuỗi thức ăn của riêng mình, vì sản phẩm phụ của từng nhóm vi sinh vật thường đóng vai trò là "thức ăn" cho các nhóm liền kề.[99]
Stromatolit là những cột trụ được tạo ra bởi các vi sinh vật trong thảm di chuyển từ dưới lên trên để tránh bị chôn vùi bởi trầm tích lắng đọng từ nước.[98] Đã có những cuộc tranh luận gay gắt về tính hợp lệ của các hóa thạch được cho là có niên đại từ trước 3 Ga,[100] phe phản đối cho rằng cái gọi là stromatolit có thể được hình thành bởi các quá trình phi sinh học.[63] Vào năm 2006, một phát hiện khác về stromatolit đã được báo cáo ở Úc tương đồng với những cái trước đó, các tảng đá ở đây có niên đại là 3,5 Ga.[101]
Trong các thảm vi sinh dưới nước hiện đại, lớp trên cùng thường có vi khuẩn lam quang hợp tạo ra môi trường giàu oxy, trong khi lớp dưới không có oxy và thường chỉ có hydro sulfide do các sinh vật sống ở đây thải ra.[99] Người ta ước tính rằng sự xuất hiện của quang hợp oxy bởi vi khuẩn trong thảm vi sinh làm tăng năng suất sinh học lên gấp 100 đến 1.000 lần. Tác nhân khử được sử dụng bởi quang hợp oxy là nước, dồi dào hơn nhiều so với các chất khử được tạo ra về mặt địa chất với điều kiện của quá trình quang hợp không oxy sớm hơn.[102] Từ thời điểm này trở đi, chính sự sống đã tạo ra nhiều tài nguyên cần thiết hơn chó chính nó so với sự dựa dẫm vào quá trình địa hóa trước đây.[103] Oxy độc hại đối với các sinh vật không thích nghi với nó, nhưng làm tăng đáng kể hiệu quả trao đổi chất của các sinh vật đã thích nghi với nó.[104][105] Oxy đã trở thành một thành phần quan trọng trong bầu khí quyển của Trái Đất vào khoảng 2,4 Ga.[106] Mặc dù sinh vật nhân chuẩn có thể đã có mặt sớm hơn nhiều,[107][108] quá trình oxy hóa của khí quyển là điều kiện tiên quyết cho sự tiến hóa của các tế bào nhân chuẩn phức tạp hơn, từ đó mà tất cả các sinh vật đa bào được tạo ra.[109] Ranh giới giữa các lớp giàu oxy và không có oxy trong thảm vi sinh sẽ di chuyển lên trên khi quá trình quang hợp ngừng lại vào buổi đêm, và sau đó đi xuống khi quá trình tiếp tục vào ngày hôm sau. Điều này sẽ tạo ra áp lực chọn lọc cho các sinh vật trong khu vực trung gian này để có được khả năng chịu đựng và sau đó là sử dụng oxy, có thể thông qua nội cộng sinh, nơi một sinh vật sống bên trong một sinh vật khác và cả hai đều được hưởng lợi từ sự hợp tác này.[23]
Vi khuẩn lam có "bộ công cụ" sinh hóa hoàn chỉnh nhất trong số tất cả các sinh vật hình thành thảm. Do đó, chúng là loài sinh vật tự cung tự cấp nhất và thích nghi tốt để tự trôi nổi dưới dạng thảm nổi và là thế hệ đầu tiên của thực vật phù du, cung cấp nền tảng cho hầu hết các chuỗi thức ăn biển.[23]
Sự đa dạng hóa của sinh vật nhân chuẩn
[sửa | sửa mã nguồn]| Eukarya |
| |||||||||||||||||||||||||||||||||
Chất nhiễm sắc, nhân, hệ thống nội màng, và ty thể
[sửa | sửa mã nguồn]Sinh vật nhân chuẩn có thể đã có mặt từ lâu trước quá trình oxy hóa khí quyển,[107] nhưng hầu hết các sinh vật nhân chuẩn hiện đại đều cần oxy, mà ty thể của chúng sử dụng để cung cấp ATP, nguồn cung cấp năng lượng bên trong của tất cả các tế bào đã biết.[109] Vào những năm 1970, có đề xuất và, sau nhiều cuộc tranh luận, đã chấp nhận rộng rãi rằng sinh vật nhân chuẩn xuất hiện do một chuỗi các sự nội cộng sinh giữa các "prokaryote". Ví dụ: một vi sinh vật săn mồi đã xâm chiếm một prokaryote lớn, có thể là một con vi khuẩn cổ, nhưng cuộc tấn công đã bị vô hiệu hóa, và kẻ tấn công đã cư trú trong nó và tiến hóa thành ty thể đầu tiên; một trong những con chimera này sau đó đã cố gắng nuốt một con vi khuẩn lam quang hợp, nhưng nạn nhân đã sống sót bên trong kẻ tấn công và sự kết hợp mới trở thành tổ tiên của thực vật; và tiếp tục. Sau khi mỗi sinh vật nội cộng sinh bắt đầu, kẻ đồng hành sẽ loại bỏ các sao chép chức năng di truyền không hiệu quả bằng cách sắp xếp lại bộ gen của chúng, một quá trình đôi khi liên quan đến việc trao đổi hệ gen giữa chúng.[112][113][114] Một giả thuyết khác cho rằng ty thể ban đầu là các endosymbiont chuyển hóa lưu huỳnh hoặc hydro, và sau đó trở thành người tiêu thụ oxy.[115] Mặt khác, ty thể có thể là một phần gốc của sinh vật nhân chuẩn.[116]
Có một cuộc tranh luận về việc khi nào sinh vật nhân chuẩn xuất hiện lần đầu tiên: sự hiện diện của steran trong tầng đá phiến ở Úc có thể chỉ ra rằng sinh vật nhân chuẩn có mặt cách đây 2.7 Ga;[108] tuy nhiên, một phân tích năm 2008 đã kết luận rằng các hóa chất này đã xâm nhập vào các tảng đá dưới 2.2 Ga và không chứng minh được gì về nguồn gốc của sinh vật nhân chuẩn.[117] Hóa thạch của loài tảo Grypania đã được báo cáo trong các tảng đá 1,85 tỷ năm tuổi (ban đầu là 2.1 Ga nhưng sau đó được sửa đổi[26]), và chỉ ra rằng sinh vật nhân chuẩn có bào quan đã phát triển.[118] Một bộ sưu tập tảo hóa thạch đa dạng đã được tìm thấy trong các loại đá có niên đại từ 1,5 đến 1,4 Ga.[119] Hóa thạch nấm được biết đến sớm nhất có niên đại từ 1.43 Ga.[120]
Lạp thể
[sửa | sửa mã nguồn]Lạp thể, siêu lớp của các bào quan trong đó lục lạp là mẫu mực nổi tiếng nhất, được cho là có nguồn gốc từ vi khuẩn lam nội bào. Sự cộng sinh tiến hóa vào khoảng 1,5 Ga và cho phép sinh vật nhân chuẩn thực hiện quá trình quang hợp oxy.[109] Kể từ đó, ba dòng tiến hóa đã xuất hiện trong đó các lạp thể được đặt tên khác nhau: lục lạp (chloroplast) trong tảo xanh và thực vật, hồng lạp (rhodoplast) trong tảo đỏ và lam tử (cyanelle hay cyanoplast) trong glaucophyte.
Sinh sản hữu tính và sinh vật đa bào
[sửa | sửa mã nguồn]Tiến hóa của sinh sản hữu tính
[sửa | sửa mã nguồn]Các đặc điểm xác định của sinh sản hữu tính ở sinh vật nhân chuẩn là giảm phân và thụ tinh. Có nhiều sự tái tổ hợp di truyền trong kiểu sinh sản này, trong đó con cái nhận được 50% gen của chúng từ bố mẹ,[121] trái ngược với kiểu sinh sản vô tính, trong đó có không có sự tái tổ hợp. Vi khuẩn trao đổi DNA theo phương pháp giao nạp vi khuẩn, lợi ích của nó bao gồm cho sinh vật khả năng kháng thuốc kháng sinh và các chất độc khác, và khả năng sử dụng các chất chuyển hóa mới.[122] Tuy nhiên, giao nạp không phải là phương tiện sinh sản và không giới hạn ở các thành viên của cùng một loài - có những trường hợp vi khuẩn chuyển DNA cho thực vật và động vật.[123]
Mặt khác, biến nạp vi khuẩn rõ ràng là sự thích nghi để chuyển DNA giữa các vi khuẩn cùng loài. Biến nạp vi khuẩn là một quá trình phức tạp liên quan đến các sản phẩm của nhiều gen vi khuẩn và có thể được coi là một dạng giao hợp của vi khuẩn.[124][125] Quá trình này xảy ra tự nhiên ở ít nhất 67 loài sinh vật nhân sơ (ở bảy ngành khác nhau).[126] Sinh sản hữu tính ở sinh vật nhân chuẩn có thể đã tiến hóa từ sự biến nạp vi khuẩn.[127]
Những nhược điểm của sinh sản hữu tính có được biết đến: sự tái tổ hợp di truyền có thể phá vỡ các tổ hợp gen có lợi và vì con đực không trực tiếp tăng số lượng con đẻ ở thế hệ tiếp theo, nên một quần thể vô tính có thể sinh sản và thay thế ít nhất là 50 thế hệ so với một quần thể hữu tính bình đẳng ở mọi khía cạnh khác.[121] Nhưng dù vậy, phần lớn động vật, thực vật, nấm và sinh vật nguyên sinh sinh sản hữu tính. Có bằng chứng mạnh mẽ cho thấy sinh sản hữu tính xuất hiện sớm trong lịch sử của sinh vật nhân chuẩn và rằng các gen kiểm soát nó thay đổi rất ít kể từ đó.[128] Làm thế nào sinh sản hữu tính phát triển và sống sót là một câu hỏi còn bỏ ngỏ.[129]

Giả thuyết Nữ hoàng Đỏ cho rằng sinh sản hữu tính giúp bảo vệ chống lại ký sinh trùng, vì ký sinh trùng dễ dàng tiến hóa hơn để vượt qua hệ đề kháng của các thế hệ vô tính giống hệt nhau về mặt di truyền so với các loài hữu tính, và có một số bằng chứng thực nghiệm cho điều này. Tuy nhiên, vẫn còn có nghi ngờ về việc liệu nó có giải thích được sự sống sót của các hữu tính hay không trong khi vẫn có nhiều loài vô tính còn tồn tại, vì một trong những dòng vô tính có thể sống sót sau các cuộc tấn công của ký sinh trùng đủ lâu để tiếp tục sinh sản.[121] Hơn nữa, trái với dự đoán của giả thuyết Nữ hoàng Đỏ, Kathryn A. Hanley et al. nhận thấy rằng tỷ lệ hiện hành, sự phong phú và cường độ trung bình của ve cao hơn đáng kể ở tắc kè tính dục so với những con vô tính cùng chung môi trường sống.[131] Ngoài ra, nhà sinh vật học Matthew Parker, sau khi xem xét nhiều nghiên cứu di truyền về tính kháng bệnh thực vật, đã không tìm thấy một ví dụ nào phù hợp với khái niệm mầm bệnh là tác nhân chọn lọc chính chịu trách nhiệm cho sinh sản hữu tính ở vật chủ.[132]
Giả thuyết đột biến xác định (DMH) của Alexey Kondrashov giả định rằng mỗi sinh vật có nhiều hơn một đột biến có hại và tác động kết hợp của các đột biến này có hại hơn so với tổng tác hại của từng đột biến riêng lẻ. Nếu vậy, sự tái tổ hợp gen sẽ làm giảm tác hại của các đột biến xấu gây ra cho thế hệ sau và đồng thời loại bỏ một số đột biến xấu từ vốn gen bằng cách phân lập chúng ở những cá thể bị diệt vong nhanh chóng vì chúng có số lượng đột biến xấu trên trung bình. Tuy nhiên, bằng chứng cho thấy các giả định của DMH rất lung lay vì nhiều loài có trung bình ít hơn một đột biến có hại cho mỗi cá thể và không có loài nào được thí nghiệm cho thấy bằng chứng về sự phối hợp giữa các đột biến có hại.[121] (Những chỉ trích tiếp theo về giả thuyết này được thảo luận trong bài viết Sự tiến hóa của sinh sản hữu tính # Loại bỏ các gen nguy hiểm)
Bản chất ngẫu nhiên của tái hợp làm cho sự phong phú tương đối của các tính trạng thay thế thay đổi từ thế hệ này sang thế hệ khác. Sự trôi dạt di truyền này không đủ để tạo ra lợi thế cho sinh sản hữu tính, nhưng sự kết hợp giữa trôi dạt di truyền và chọn lọc tự nhiên có thể là đủ. Khi cơ hội tạo ra sự kết hợp của các tính trạng tốt, chọn lọc tự nhiên mang lại lợi thế lớn cho các dòng dõi trong đó các tính trạng này trở thành liên kết di truyền. Mặt khác, lợi ích của các tính trạng tốt bị trung hòa nếu chúng xuất hiện cùng với các tính trạng xấu. Tái hợp tính dục mang lại những đặc điểm tốt cho các cơ hội được liên kết với các đặc điểm tốt khác và các mô hình toán học cho thấy điều này có thể là quá đủ để bù đắp những nhược điểm của sinh sản hữu tính.[129] Các giả thuyết khác không thỏa đáng cũng đang được xem xét.[121]
Các giả thuyết dưới đây có thể giải thích tại sao và bằng cách nào giao phối lại tiến hóa:
- Nó cho phép các sinh vật sửa chữa các lỗi về gen.[133]
Chức năng thích ứng của giới tính ngày nay vẫn là một vấn đề lớn chưa được giải quyết trong sinh học. Các mô hình cạnh tranh để giải thích chức năng thích nghi của giới tính đã được xem xét bởi John A. Birdsell và Christopher Wills.[134] Các giả thuyết được thảo luận ở trên đều phụ thuộc vào các tác động có lợi khả thi của biến dị di truyền ngẫu nhiên được tạo ra bởi sự tái hợp di truyền. Một quan điểm khác là giới tính nảy sinh và được duy trì, như là một quá trình sửa chữa tổn thương DNA và biến thể di truyền được tạo ra là một sản phẩm phụ đôi khi có lợi.[127][135]
Tính đa bào
[sửa | sửa mã nguồn]Các định nghĩa đơn giản nhất về "đa bào", ví dụ "có nhiều tế bào", có thể bao gồm các tập đoàn vi khuẩn lam như Nostoc. Ngay cả một định nghĩa học thuật như "có cùng bộ gen nhưng các loại tế bào khác nhau" vẫn sẽ bao gồm một số chi của Volvox tảo xanh, có các tế bào chuyên sinh sản.[136] Đa bào phát triển độc lập trong các sinh vật đa dạng như bọt biển và các động vật khác, nấm, thực vật, tảo nâu, vi khuẩn lam, nấm mốc và vi khuẩn myxobacteria.[26][137] Để đơn giản, bài viết này tập trung vào các sinh vật thể hiện sự chuyên hóa lớn nhất trong các tế bào và sở hữu nhiều loại tế bào khác nhau, mặc dù cách tiếp cận tiến hóa phức tạp sinh học này có thể được coi là "khá duy nhân."[27]
Những lợi thế ban đầu của đa bào có thể bao gồm: hấp thụ hiệu quả hơn các chất dinh dưỡng được tiêu hóa bên ngoài tế bào,[138] tăng sức phản kháng lại động vật ăn thịt, nhiều trong số đó bị tấn công bằng cách nhấn chìm; khả năng chống lại dòng chảy bằng cách gắn vào một bề mặt vững chắc; khả năng tiếp cận lên bề mặt để lọc thức ăn hoặc thu được ánh sáng mặt trời phục vụ quang hợp;[139]] khả năng tạo môi trường bên trong giúp bảo vệ chống lại môi trường bên ngoài;[27] và cả cơ hội cho một nhóm tế bào tỏ ra "thông minh" bằng cách chia sẻ thông tin.[140] Những đặc điểm này cũng sẽ tạo cơ hội cho các sinh vật khác đa dạng hóa, bằng cách tạo ra nhiều môi trường phong phú hơn so với các thảm vi sinh có thể.[139]
Tính đa bào của các tế bào biệt hóa có lợi cho toàn bộ sinh vật nhưng bất lợi đôi với từng tế bào, hầu hết đều mất cơ hội tự sinh sản. Trong một sinh vật đa bào vô tính, các tế bào bất hảo còn giữ khả năng sinh sản có thể tiếp quản và làm giảm sinh vật xuống một khối các tế bào không biệt hóa. Sinh sản hữu tính giúp loại bỏ các tế bào bất hảo như vậy từ thế hệ tiếp theo và do đó dường như là điều kiện tiên quyết cho tính đa bào phức tạp.[139]
Bằng chứng sẵn có chỉ ra rằng sinh vật nhân chuẩn tiến hóa sớm hơn nhiều nhưng vẫn không rõ ràng cho đến khi đa dạng hóa nhanh chóng vào khoảng 1 Ga. Khía cạnh duy nhất trong đó sinh vật nhân chuẩn rõ ràng vượt trội hơn vi khuẩn và vi khuẩn cổ là sự đa dạng kiểu hình, và khả năng sinh sản giúp chúng lợi dụng điều này và tạo ra nhiều hình dạng với các chức năng khác nhau.[139]
Tham khảo
[sửa | sửa mã nguồn]- ^ a b Pearce, Ben K.D.; Tupper, Andrew S.; Pudritz, Ralph E.; Higgs, Paul G. (ngày 1 tháng 3 năm 2018). "Constraining the Time Interval for the Origin of Life on Earth". Astrobiology (bằng tiếng Anh). Quyển 18 số 3. tr. 343–364. Bibcode:2018AsBio..18..343P. doi:10.1089/ast.2017.1674. PMID 29570409.
- ^ Rosing, Minik T. (ngày 29 tháng 1 năm 1999). "13C-Depleted Carbon Microparticles in >3700-Ma Sea-Floor Sedimentary Rocks from West Greenland". Science (bằng tiếng Anh). Quyển 283 số 5402. tr. 674–676. Bibcode:1999Sci...283..674R. doi:10.1126/science.283.5402.674. ISSN 0036-8075. PMID 9924024.
- ^ a b Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi; Nagase, Toshiro; Rosing, Minik T. (tháng 1 năm 2014). "Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks". Nature Geoscience (bằng tiếng Anh). Quyển 7 số 1. tr. 25–28. Bibcode:2014NatGe...7...25O. doi:10.1038/ngeo2025. ISSN 1752-0908.
- ^ Papineau, D.; Gregorio, B. T. De; Cody, G. D.; O’Neil, J.; Steele, A.; Stroud, R. M.; Fogel, M. L. (tháng 6 năm 2011). "Young poorly crystalline graphite in the >3.8-Gyr-old Nuvvuagittuq banded iron formation". Nature Geoscience (bằng tiếng Anh). Quyển 4 số 6. tr. 376–379. Bibcode:2011NatGe...4..376P. doi:10.1038/ngeo1155. ISSN 1752-0908.
- ^ "Life on Earth likely started 4.1 billion years ago—much earlier than scientists thought". ngày 19 tháng 10 năm 2015.
- ^ Bell, Elizabeth A.; Boehnke, Patrick; Harrison, T. Mark; Mao, Wendy L. (ngày 24 tháng 11 năm 2015). "Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon". Proceedings of the National Academy of Sciences. Quyển 112 số 47. tr. 14518–14521. Bibcode:2015PNAS..11214518B. doi:10.1073/pnas.1517557112. PMC 4664351. PMID 26483481.
- ^ Nemchin, Alexander A.; Whitehouse, Martin J.; Menneken, Martina; Geisler, Thorsten; Pidgeon, Robert T.; Wilde, Simon A. (tháng 7 năm 2008). "A light carbon reservoir recorded in zircon-hosted diamond from the Jack Hills". Nature (bằng tiếng Anh). Quyển 454 số 7200. tr. 92–95. Bibcode:2008Natur.454...92N. doi:10.1038/nature07102. ISSN 1476-4687. PMID 18596808.
- ^ Futuyma 2005
- ^ McKinney 1997, tr. 110
- ^ Stearns, Beverly Peterson; Stearns, S. C.; Stearns, Stephen C. (2000). Watching, from the Edge of Extinction. Yale University Press. tr. reface x. ISBN 978-0-300-08469-6. Truy cập ngày 30 tháng 5 năm 2017.
- ^ Novacek, Michael J. (ngày 8 tháng 11 năm 2014). "Prehistory's Brilliant Future". The New York Times. New York: The New York Times Company. Truy cập ngày 25 tháng 12 năm 2014.
- ^ Mora, Camilo; Tittensor, Derek P.; Adl, Sina; và đồng nghiệp (ngày 23 tháng 8 năm 2011). "How Many Species Are There on Earth and in the Ocean?". PLOS Biology. Quyển 9 số 8. tr. e1001127. doi:10.1371/journal.pbio.1001127. PMC 3160336. PMID 21886479.
{{Chú thích tạp chí}}: Quản lý CS1: DOI truy cập mở nhưng không được đánh ký hiệu (liên kết) - ^ Miller & Spoolman 2012, tr. 62
- ^ Chapman, Arthur D. (2009). Numbers of Living Species in Australia and the World. 2nd edition. Canberra: Australian Biological Resources Study. ISBN 978-0-642-56860-1. Truy cập ngày 6 tháng 11 năm 2016.
- ^ "Catalogue of Life: 2016 Annual Checklist". 2016. Truy cập ngày 6 tháng 11 năm 2016.
- ^ Staff (ngày 2 tháng 5 năm 2016). "Researchers find that Earth may be home to 1 trillion species". National Science Foundation. Truy cập ngày 11 tháng 12 năm 2016.
- ^ "Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures".
- ^ a b Borenstein, Seth (ngày 19 tháng 10 năm 2015). "Hints of life on what was thought to be desolate early Earth". Excite. Yonkers, NY: Mindspark Interactive Network. Associated Press. Bản gốc lưu trữ ngày 23 tháng 10 năm 2015. Truy cập ngày 8 tháng 10 năm 2018.
{{Chú thích báo}}: Đã định rõ hơn một tham số trong|archiveurl=và|archive-url=(trợ giúp) - ^ Bell, Elizabeth A.; Boehnike, Patrick; Harrison, T. Mark; và đồng nghiệp (ngày 19 tháng 10 năm 2015). "Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon" (PDF). Proc. Natl. Acad. Sci. U.S.A. Quyển 112 số 47. tr. 14518–21. Bibcode:2015PNAS..11214518B. doi:10.1073/pnas.1517557112. PMC 4664351. PMID 26483481. Truy cập ngày 20 tháng 10 năm 2015. Early edition, published online before print.
- ^ a b Dodd, Matthew S.; Papineau, Dominic; Grenne, Tor; slack, John F.; Rittner, Martin; Pirajno, Franco; O'Neil, Jonathan; Little, Crispin T. S. (ngày 2 tháng 3 năm 2017). "Evidence for early life in Earth's oldest hydrothermal vent precipitates". Nature. Quyển 543 số 7643. tr. 60–64. Bibcode:2017Natur.543...60D. doi:10.1038/nature21377. Bản gốc lưu trữ ngày 12 tháng 9 năm 2018. Truy cập ngày 16 tháng 12 năm 2017.
- ^ "Scientists Say Canadian Bacteria Fossils May Be Earth's Oldest".
- ^ Ghosh, Pallab (ngày 1 tháng 3 năm 2017). "Earliest evidence of life on Earth 'found'". BBC News. Truy cập ngày 2 tháng 3 năm 2017.
- ^ a b c Nisbet, Euan G.; Fowler, C. M. R. (ngày 7 tháng 12 năm 1999). "Archaean metabolic evolution of microbial mats". Proceedings of the Royal Society B. Quyển 266 số 1436. tr. 2375–2382. doi:10.1098/rspb.1999.0934. PMC 1690475.
- ^ Anbar, Ariel D.; Yun Duan; Lyons, Timothy W.; và đồng nghiệp (ngày 28 tháng 9 năm 2007). "A Whiff of Oxygen Before the Great Oxidation Event?". Science. Quyển 317 số 5846. tr. 1903–1906. Bibcode:2007Sci...317.1903A. doi:10.1126/science.1140325. PMID 17901330.
- ^ Knoll, Andrew H.; Javaux, Emmanuelle J.; Hewitt, David; Cohen, Phoebe (ngày 29 tháng 6 năm 2006). "Eukaryotic organisms in Proterozoic oceans". Philosophical Transactions of the Royal Society B. Quyển 361 số 1470. tr. 1023–1038. doi:10.1098/rstb.2006.1843. PMC 1578724. PMID 16754612.
- ^ a b c d e Fedonkin, Mikhail A. (ngày 31 tháng 3 năm 2003). "The origin of the Metazoa in the light of the Proterozoic fossil record" (PDF). Paleontological Research. Quyển 7 số 1. tr. 9–41. doi:10.2517/prpsj.7.9. Bản gốc (PDF) lưu trữ ngày 26 tháng 2 năm 2009. Truy cập ngày 2 tháng 9 năm 2008.
- ^ a b c Bonner, John Tyler (1998). "The origins of multicellularity". Integrative Biology. Quyển 1 số 1. tr. 27–36. doi:10.1002/(SICI)1520-6602(1998)1:1<27::AID-INBI4>3.0.CO;2-6.
- ^ Otto, Sarah P.; Lenormand, Thomas (tháng 4 năm 2002). "Evolution of sex: Resolving the paradox of sex and recombination". Nature Reviews Genetics. Quyển 3 số 4. tr. 252–261. doi:10.1038/nrg761. PMID 11967550.
- ^ Letunic, Ivica; Bork, Peer. "iTOL: Interactive Tree of Life". Heidelberg, Germany: European Molecular Biology Laboratory. Truy cập ngày 21 tháng 7 năm 2015.
- ^ Fedonkin, Mikhail A.; Simonetta, Alberto; Ivantsov, Andrei Yu. (2007). "New data on Kimberella, the Vendian mollusc-like organism (White Sea region, Russia): palaeoecological and evolutionary implications" (PDF). Geological Society Special Publication. Quyển 286. tr. 157–179. Bibcode:2007GSLSP.286..157F. doi:10.1144/SP286.12. Truy cập ngày 16 tháng 5 năm 2013.
- ^ "The late Precambrian greening of the Earth". nature.com. Truy cập ngày 10 tháng 4 năm 2018.
- ^ Bản mẫu:Url= http://www.nature.com/articles/nature09943
- ^ Beraldi-Campesi, Hugo (ngày 23 tháng 2 năm 2013). "Early life on land and the first terrestrial ecosystems". Ecological Processes. Quyển 2 số 1. tr. 4. doi:10.1186/2192-1709-2-1. Truy cập ngày 2 tháng 9 năm 2017.
{{Chú thích tạp chí}}: Quản lý CS1: DOI truy cập mở nhưng không được đánh ký hiệu (liên kết) - ^ Algeo, Thomas J.; Scheckler, Stephen E. (ngày 29 tháng 1 năm 1998). "Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events". Philosophical Transactions of the Royal Society B. Quyển 353 số 1365. tr. 113–130. doi:10.1098/rstb.1998.0195. PMC 1692181.
- ^ Jun-Yuan Chen; Oliveri, Paola; Chia-Wei Li; và đồng nghiệp (ngày 25 tháng 4 năm 2000). "Precambrian animal diversity: Putative phosphatized embryos from the Doushantuo Formation of China". Proc. Natl. Acad. Sci. U.S.A. Quyển 97 số 9. tr. 4457–4462. Bibcode:2000PNAS...97.4457C. doi:10.1073/pnas.97.9.4457. PMC 18256. PMID 10781044. Truy cập ngày 30 tháng 4 năm 2009.
- ^ D-G. Shu; H-L. Luo; Conway Morris, Simon; và đồng nghiệp (ngày 4 tháng 11 năm 1999). "Lower Cambrian vertebrates from south China" (PDF). Nature. Quyển 402 số 6757. tr. 42–46. Bibcode:1999Natur.402...42S. doi:10.1038/46965. Bản gốc (PDF) lưu trữ ngày 26 tháng 2 năm 2009. Truy cập ngày 22 tháng 1 năm 2015.
- ^ Hoyt, Donald F. (ngày 17 tháng 2 năm 1997). "Synapsid Reptiles". ZOO 138 Vertebrate Zoology (Lecture). Pomona, CA: California State Polytechnic University, Pomona. Bản gốc lưu trữ ngày 20 tháng 5 năm 2009. Truy cập ngày 22 tháng 1 năm 2015.
- ^ Barry, Patrick L. (ngày 28 tháng 1 năm 2002). Phillips, Tony (biên tập). "The Great Dying". Science@NASA. Science and Technology Directorate, Marshall Space Flight Center. Truy cập ngày 22 tháng 1 năm 2015.
- ^ Tanner, Lawrence H.; Lucas, Spencer G.; Chapman, Mary G. (tháng 3 năm 2004). "Assessing the record and causes of Late Triassic extinctions" (PDF). Earth-Science Reviews. Quyển 65 số 1–2. tr. 103–139. Bibcode:2004ESRv...65..103T. doi:10.1016/S0012-8252(03)00082-5. Bản gốc (PDF) lưu trữ ngày 25 tháng 10 năm 2007. Truy cập ngày 22 tháng 10 năm 2007.
- ^ Benton 1997
- ^ Fastovsky, David E.; Sheehan, Peter M. (tháng 3 năm 2005). "The Extinction of the Dinosaurs in North America" (PDF). GSA Today. Quyển 15 số 3. tr. 4–10. doi:10.1130/1052-5173(2005)015<4:TEOTDI>2.0.CO;2. Bản gốc (PDF) lưu trữ ngày 9 tháng 8 năm 2017. Truy cập ngày 23 tháng 1 năm 2015.
- ^ Roach, John (ngày 20 tháng 6 năm 2007). "Dinosaur Extinction Spurred Rise of Modern Mammals". ZOO 138 Vertebrate Zoology (Lecture). Washington, D.C.: National Geographic Society. Bản gốc lưu trữ ngày 20 tháng 5 năm 2009. Truy cập ngày 8 tháng 3 năm 2009.
- ^ 252 Ma
- ^ Erwin, Douglas H. (ngày 9 tháng 11 năm 2015). "Early metazoan life: divergence, environment and ecology". Phil. Trans. R. Soc. B. Quyển 370 số 20150036. tr. 20150036. doi:10.1098/rstb.2015.0036. PMC 4650120. PMID 26554036.
- ^ El Albani, Abderrazak; Bengtson, Stefan; Canfield, Donald E.; và đồng nghiệp (ngày 1 tháng 7 năm 2010). "Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago". Nature. Quyển 466 số 7302. tr. 100–104. Bibcode:2010Natur.466..100A. doi:10.1038/nature09166. PMID 20596019.
- ^ Dalrymple 1991
- Newman 2007
- Dalrymple, G. Brent (2001). "The age of the Earth in the twentieth century: a problem (mostly) solved". Geological Society Special Publication. Quyển 190 số 1. tr. 205–221. Bibcode:2001GSLSP.190..205D. doi:10.1144/GSL.SP.2001.190.01.14. Truy cập ngày 23 tháng 1 năm 2015.
- ^ Galimov, Erik M.; Krivtsov, Anton M. (tháng 12 năm 2005). "Origin of the Earth—Moon system" (PDF). Journal of Earth System Science. Quyển 114 số 6. tr. 593–600. Bibcode:2005JESS..114..593G. doi:10.1007/BF02715942. Truy cập ngày 23 tháng 1 năm 2015.
- ^ Thompson, Andrea (ngày 25 tháng 9 năm 2008). "Oldest Rocks on Earth Found". LiveScience. Watsonville, CA: Imaginova. Truy cập ngày 23 tháng 1 năm 2015.
- ^ Dalrymple 1991
- ^ Newman 2007
- ^ Dalrymple, G. Brent (2001). "The age of the Earth in the twentieth century: a problem (mostly) solved". Geological Society Special Publication. Quyển 190. tr. 205–221. Bibcode:2001GSLSP.190..205D. doi:10.1144/GSL.SP.2001.190.01.14. Truy cập ngày 23 tháng 1 năm 2015.
- ^ Cohen, Barbara A.; Swindle, Timothy D.; Kring, David A. (ngày 1 tháng 12 năm 2000). "Support for the Lunar Cataclysm Hypothesis from Lunar Meteorite Impact Melt Ages". Science. Quyển 290 số 5497. tr. 1754–1756. Bibcode:2000Sci...290.1754C. doi:10.1126/science.290.5497.1754. PMID 11099411. Truy cập ngày 23 tháng 1 năm 2015.
- ^ "Early Earth Likely Had Continents And Was Habitable" (Thông cáo báo chí). Boulder, CO: University of Colorado. ngày 17 tháng 11 năm 2005. Bản gốc lưu trữ ngày 24 tháng 1 năm 2015. Truy cập ngày 23 tháng 1 năm 2015.
- ^ Cavosie, Aaron J.; Valley, John W.; Wilde, Simon A.; Edinburgh Ion Microprobe Facility (ngày 15 tháng 7 năm 2005). "Magmatic δ18O in 4400–3900 Ma detrital zircons: A record of the alteration and recycling of crust in the Early Archean". Earth and Planetary Science Letters. Quyển 235 số 3–4. tr. 663–681. Bibcode:2005E&PSL.235..663C. doi:10.1016/j.epsl.2005.04.028.
- ^ a b c Garwood, Russell J. (2012). "Patterns In Palaeontology: The first 3 billion years of evolution". Palaeontology Online. Quyển 2 số 11. tr. 1–14. Bản gốc lưu trữ ngày 26 tháng 6 năm 2015. Truy cập ngày 25 tháng 6 năm 2015.
- ^ Britt, Robert Roy (ngày 24 tháng 7 năm 2002). "Evidence for Ancient Bombardment of Earth". Space.com. New York: Space Holding Corp. Bản gốc lưu trữ ngày 4 tháng 10 năm 2002. Truy cập ngày 23 tháng 1 năm 2015.
{{Chú thích báo}}: Đã định rõ hơn một tham số trong|archiveurl=và|archive-url=(trợ giúp) - ^ Valley, John W.; Peck, William H.; King, Elizabeth M.; Wilde, Simon A. (tháng 4 năm 2002). "A cool early Earth" (PDF). Geology. Quyển 30 số 4. tr. 351–354. Bibcode:2002Geo....30..351V. doi:10.1130/0091-7613(2002)030<0351:ACEE>2.0.CO;2. Truy cập ngày 13 tháng 9 năm 2008.
{{Chú thích tạp chí}}: Quản lý CS1: ngày tháng và năm (liên kết)CS1 maint: Date and year (link) Valley, John W.; Peck, William H.; King, Elizabeth M.; Wilde, Simon A. (tháng 4 năm 2002). "A cool early Earth" (PDF). Geology. Quyển 30 số 4. tr. 351–354. Bibcode:2002Geo....30..351V. doi:10.1130/0091-7613(2002)030<0351:ACEE>2.0.CO;2. Truy cập ngày 13 tháng 9 năm 2008.{{Chú thích tạp chí}}: Quản lý CS1: ngày tháng và năm (liên kết) - ^ Dauphas, Nicolas; Robert, François; Marty, Bernard (tháng 12 năm 2000). "The Late Asteroidal and Cometary Bombardment of Earth as Recorded in Water Deuterium to Protium Ratio". Icarus. Quyển 148 số 2. tr. 508–512. Bibcode:2000Icar..148..508D. doi:10.1006/icar.2000.6489.
- ^ Scalice, Daniella (ngày 20 tháng 5 năm 2009). Fletcher, Julie (biên tập). "Microbial Habitability During the Late Heavy Bombardment". Astrobiology. Mountain View, CA: NASA Astrobiology Program. Bản gốc lưu trữ ngày 24 tháng 1 năm 2015. Truy cập ngày 23 tháng 1 năm 2015.
- ^ a b Brasier, Martin; McLoughlin, Nicola; Green, Owen; Wacey, David (tháng 6 năm 2006). "A fresh look at the fossil evidence for early Archaean cellular life" (PDF). Philosophical Transactions of the Royal Society B. Quyển 361 số 1470. tr. 887–902. doi:10.1098/rstb.2006.1835. PMC 1578727. PMID 16754605. Truy cập ngày 30 tháng 8 năm 2008.
- ^ Schopf, J. William (ngày 30 tháng 4 năm 1993). "Microfossils of the Early Archean Apex Chert: New Evidence of the Antiquity of Life". Science. Quyển 260 số 5108. tr. 640–646. Bibcode:1993Sci...260..640S. doi:10.1126/science.260.5108.640. PMID 11539831. Truy cập ngày 24 tháng 1 năm 2015.
- ^ Mojzsis, Stephen J.; Arrhenius, Gustaf; McKeegan, Kevin D.; và đồng nghiệp (tháng 11 năm 1996). "Evidence for life on Earth before 3,800 million years ago". Nature. Quyển 384 số 6604. tr. 55–59. Bibcode:1996Natur.384...55M. doi:10.1038/384055a0. PMID 8900275. Truy cập ngày 30 tháng 8 năm 2008.
- ^ a b Grotzinger, John P.; Rothman, Daniel H. (ngày 3 tháng 10 năm 1996). "An abiotic model for stromatolite morphogenesis". Nature. Quyển 383 số 6599. tr. 423–425. Bibcode:1996Natur.383..423G. doi:10.1038/383423a0.
- ^ Fedo, Christopher M.; Whitehouse, Martin J. (ngày 24 tháng 5 năm 2002). "Metasomatic Origin of Quartz-Pyroxene Rock, Akilia, Greenland, and Implications for Earth's Earliest Life". Science. Quyển 296 số 5572. tr. 1448–1452. Bibcode:2002Sci...296.1448F. doi:10.1126/science.1070336. PMID 12029129. Truy cập ngày 24 tháng 1 năm 2015.
- Lepland, Aivo; van Zuilen, Mark A.; Arrhenius, Gustaf; và đồng nghiệp (tháng 1 năm 2005). "Questioning the evidence for Earth's earliest life—Akilia revisited". Geology. Quyển 33 số 1. tr. 77–79. Bibcode:2005Geo....33...77L. doi:10.1130/G20890.1. Truy cập ngày 24 tháng 1 năm 2015.
- ^ Schopf, J. William (ngày 29 tháng 6 năm 2006). "Fossil evidence of Archaean life". Philosophical Transactions of the Royal Society B. Quyển 361 số 1470. tr. 869–885. doi:10.1098/rstb.2006.1834. PMC 1578735. PMID 16754604.
- ^ "This May Be the Oldest Known Sign of Life on Earth". ngày 1 tháng 3 năm 2017. Truy cập ngày 2 tháng 3 năm 2017.
- ^ Ciccarelli, Francesca D.; Doerks, Tobias; von Mering, Christian; và đồng nghiệp (ngày 3 tháng 3 năm 2006). "Toward Automatic Reconstruction of a Highly Resolved Tree of Life" (Submitted manuscript). Science. Quyển 311 số 5765. tr. 1283–1287. Bibcode:2006Sci...311.1283C. CiteSeerX 10.1.1.381.9514. doi:10.1126/science.1123061. PMID 16513982.
- ^ Mason, Stephen F. (1984). "Origins of biomolecular handedness". Nature. Quyển 311 số 5981. tr. 19–23. Bibcode:1984Natur.311...19M. doi:10.1038/311019a0. PMID 6472461.
- ^ Orgel, Leslie E. (tháng 10 năm 1994). "The Origin of Life on the Earth". Scientific American. Quyển 271 số 4. tr. 76–83. Bibcode:1994SciAm.271d..76O. doi:10.1038/scientificamerican1094-76. PMID 7524147. Bản gốc (PDF) lưu trữ ngày 24 tháng 1 năm 2001. Truy cập ngày 30 tháng 8 năm 2008.
- ^ a b Bennett 2008, tr. 82–85
- ^ Schulze-Makuch, Dirk; Irwin, Louis N. (tháng 4 năm 2006). "The prospect of alien life in exotic forms on other worlds". Naturwissenschaften. Quyển 93 số 4. tr. 155–72. Bibcode:2006NW.....93..155S. doi:10.1007/s00114-005-0078-6. PMID 16525788.
- ^ Peretó, Juli (2005). "Controversies on the origin of life" (PDF). International Microbiology. Quyển 8 số 1. tr. 23–31. PMID 15906258. Bản gốc (PDF) lưu trữ ngày 24 tháng 8 năm 2015. Truy cập ngày 7 tháng 10 năm 2007.
- ^ Szathmáry, Eörs (ngày 3 tháng 2 năm 2005). "Life: In search of the simplest cell". Nature. Quyển 433 số 7025. tr. 469–470. Bibcode:2005Natur.433..469S. doi:10.1038/433469a. PMID 15690023. Truy cập ngày 1 tháng 9 năm 2008.
- ^ Luisi, Pier Luigi; Ferri, Francesca; Stano, Pasquale (tháng 1 năm 2006). "Approaches to semi-synthetic minimal cells: a review". Naturwissenschaften. Quyển 93 số 1. tr. 1–13. Bibcode:2006NW.....93....1L. doi:10.1007/s00114-005-0056-z. PMID 16292523.
- ^ Hoenigsberg, Hugo (ngày 30 tháng 12 năm 2003). "Evolution without speciation but with selection: LUCA, the Last Universal Common Ancestor in Gilbert's RNA world". Genetics and Molecular Research. Quyển 2 số 4. tr. 366–375. PMID 15011140. Truy cập ngày 30 tháng 8 năm 2008.
- ^ a b Trevors, Jack T.; Abel, David L. (tháng 11 năm 2004). "Chance and necessity do not explain the origin of life". Cell Biology International. Quyển 28 số 11. tr. 729–739. doi:10.1016/j.cellbi.2004.06.006. PMID 15563395.
- ^ a b c Forterre, Patrick; Benachenhou-Lahfa, Nadia; Confalonieri, Fabrice; và đồng nghiệp (1992). "The nature of the last universal ancestor and the root of the tree of life, still open questions". BioSystems. Quyển 28 số 1–3. tr. 15–32. doi:10.1016/0303-2647(92)90004-I. PMID 1337989.
- ^ a b Cech, Thomas R. (ngày 11 tháng 8 năm 2000). "The Ribosome Is a Ribozyme". Science. Quyển 289 số 5481. tr. 878–879. doi:10.1126/science.289.5481.878. PMID 10960319. Truy cập ngày 26 tháng 1 năm 2015.
- ^ Pearce, Ben K. D.; Pudritz, Ralph E.; Semenov, Dmitry A.; Henning, Thomas K. (ngày 24 tháng 10 năm 2017). "Origin of the RNA world: The fate of nucleobases in warm little ponds". Proceedings of the National Academy of Sciences (bằng tiếng Anh). Quyển 114 số 43. tr. 11327–11332. doi:10.1073/pnas.1710339114. ISSN 0027-8424. PMID 28973920.
- ^ Pearce, Ben K. D.; Pudritz, Ralph E.; Semenov, Dmitry A.; Henning, Thomas K. (ngày 24 tháng 10 năm 2017). "Origin of the RNA world: The fate of nucleobases in warm little ponds". Proceedings of the National Academy of Sciences (bằng tiếng Anh). Quyển 114 số 43. tr. 11327–11332. Bibcode:2017PNAS..11411327P. doi:10.1073/pnas.1710339114. ISSN 0027-8424. PMC 5664528. PMID 28973920.
- ^ Martin, William; Russell, Michael J. (ngày 29 tháng 1 năm 2003). "On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells". Philosophical Transactions of the Royal Society B. Quyển 358 số 1429. tr. 59–85. doi:10.1098/rstb.2002.1183. PMC 1693102. PMID 12594918.
- ^ Wächtershäuser, Günter (ngày 25 tháng 8 năm 2000). "Life as We Don't Know It". Science. Quyển 289 số 5483. tr. 1307–1308. doi:10.1126/science.289.5483.1307. PMID 10979855.
- ^ Trevors, Jack T.; Psenner, Roland (tháng 12 năm 2001). "From self-assembly of life to present-day bacteria: a possible role for nanocells". FEMS Microbiology Reviews. Quyển 25 số 5. tr. 573–582. doi:10.1111/j.1574-6976.2001.tb00592.x. PMID 11742692.
- ^ Segré, Daniel; Ben-Eli, Dafna; Deamer, David W.; Lancet, Doron (tháng 2 năm 2001). "The Lipid World" (PDF). Origins of Life and Evolution of Biospheres. Quyển 31 số 1–2. tr. 119–145. doi:10.1023/A:1006746807104. PMID 11296516. Truy cập ngày 26 tháng 1 năm 2015.
- ^ Cairns-Smith 1968, tr. 57–66
- ^ Ferris, James P. (tháng 6 năm 1999). "Prebiotic Synthesis on Minerals: Bridging the Prebiotic and RNA Worlds". The Biological Bulletin. Quyển 196 số 3. tr. 311–314. doi:10.2307/1542957. JSTOR 1542957. PMID 10390828. "This paper was originally presented at a workshop titled Evolution: A Molecular Point of View."
- ^ Hanczyc, Martin M.; Fujikawa, Shelly M.; Szostak, Jack W. (ngày 24 tháng 10 năm 2003). "Experimental Models of Primitive Cellular Compartments: Encapsulation, Growth, and Division". Science. Quyển 302 số 5645. tr. 618–622. Bibcode:2003Sci...302..618H. doi:10.1126/science.1089904. PMC 4484575. PMID 14576428. Truy cập ngày 26 tháng 1 năm 2015.
- ^ Hartman, Hyman (tháng 10 năm 1998). "Photosynthesis and the Origin of Life". Origins of Life and Evolution of Biospheres. Quyển 28 số 4–6. tr. 512–521. Bibcode:1998OLEB...28..515H. doi:10.1023/A:1006548904157. PMID 11536891.
- ^ O'Leary 2008
- ^ a b Arrhenius 1980, tr. 32
- ^ Hoyle, Fred; Wickramasinghe, Nalin C. (tháng 11 năm 1979). "On the Nature of Interstellar Grains". Astrophysics and Space Science. Quyển 66 số 1. tr. 77–90. Bibcode:1979Ap&SS..66...77H. doi:10.1007/BF00648361.
- ^ a b Crick, Francis H.; Orgel, Leslie E (tháng 7 năm 1973). "Directed Panspermia". Icarus. Quyển 19 số 3. tr. 341–348. Bibcode:1973Icar...19..341C. doi:10.1016/0019-1035(73)90110-3.
- ^ a b c Warmflash, David; Weiss, Benjamin (tháng 11 năm 2005). "Did Life Come From Another World?". Scientific American. Quyển 293 số 5. tr. 64–71. Bibcode:2005SciAm.293e..64W. doi:10.1038/scientificamerican1105-64. PMID 16318028. Truy cập ngày 25 tháng 1 năm 2015.
- ^ Wickramasinghe, Nalin C.; Wickramasinghe, Janaki T. (tháng 9 năm 2008). "On the possibility of microbiota transfer from Venus to Earth". Astrophysics and Space Science. Quyển 317 số 1–2. tr. 133–137. Bibcode:2008Ap&SS.317..133W. CiteSeerX 10.1.1.470.2347. doi:10.1007/s10509-008-9851-2.
- ^ Clancy, Brack & Horneck 2005
- ^ Horneck, Gerda; Klaus, David M.; Mancinelli, Rocco L. (tháng 3 năm 2010). "Space Microbiology". Microbiology and Molecular Biology Reviews. Quyển 74 số 1. tr. 121–156. doi:10.1128/mmbr.00016-09. PMC 2832349. PMID 20197502. Truy cập ngày 29 tháng 7 năm 2013.
- ^ Than, Ker (ngày 23 tháng 8 năm 2007). "Claim of Martian Life Called 'Bogus'". Space.com. Watsonville, CA: Imaginova. Truy cập ngày 25 tháng 1 năm 2015.
- ^ a b Krumbein và đồng nghiệp 2003, tr. 1–28
- ^ a b Risatti, J. Bruno; Capman, William C.; Stahl, David A. (ngày 11 tháng 10 năm 1994). "Community structure of a microbial mat: The phylogenetic dimension". Proc. Natl. Acad. Sci. U.S.A. Quyển 91 số 21. tr. 10173–10177. Bibcode:1994PNAS...9110173R. doi:10.1073/pnas.91.21.10173. PMC 44980. PMID 7937858.
- ^ "Biodiversity rocks". Nature (Editor's summary). Quyển 441 số 7094. ngày 8 tháng 6 năm 2006. Truy cập ngày 10 tháng 1 năm 2009.
- ^ Allwood, Abigail C.; Walter, Malcolm R.; Kamber, Balz S.; và đồng nghiệp (ngày 8 tháng 6 năm 2006). "Stromatolite reef from the Early Archaean era of Australia". Nature. Quyển 441 số 7094. tr. 714–718. Bibcode:2006Natur.441..714A. doi:10.1038/nature04764. PMID 16760969.
- ^ Blankenship, Robert E. (ngày 1 tháng 1 năm 2001). "Molecular evidence for the evolution of photosynthesis". Trends in Plant Science. Quyển 6 số 1. tr. 4–6. doi:10.1016/S1360-1385(00)01831-8. PMID 11164357.
- ^ Hoehler, Tori M.; Bebout, Brad M.; Des Marais, David J. (ngày 19 tháng 7 năm 2001). "The role of microbial mats in the production of reduced gases on the early Earth". Nature. Quyển 412 số 6844. tr. 324–327. Bibcode:2001Natur.412..324H. doi:10.1038/35085554. PMID 11460161.
- ^ Abele, Doris (ngày 7 tháng 11 năm 2002). "Toxic oxygen: The radical life-giver" (PDF). Nature. Quyển 420 số 6911. tr. 27. Bibcode:2002Natur.420...27A. doi:10.1038/420027a. PMID 12422197.
- ^ Westerdahl, Becky B. (2007). "Introduction to Aerobic Respiration". Biological Science 10V (Lecture). Davis, CA: University of California, Davis. Bản gốc lưu trữ ngày 29 tháng 10 năm 2007. Truy cập ngày 14 tháng 7 năm 2008.
- Textbook used for lecture: Biology Today and Tomorrow With Physiology (2007), ISBN 0-495-01654-3.
- ^ Goldblatt, Colin; Lenton, Timothy M.; Watson, Andrew J. (2006). "The Great Oxidation at ~2.4 Ga as a bistability in atmospheric oxygen due to UV shielding by ozone" (PDF). Geophysical Research Abstracts. Quyển 8 số 770. SRef-ID: 1607-7962/gra/EGU06-A-00770. Bản gốc (PDF) lưu trữ ngày 8 tháng 5 năm 2019. Truy cập ngày 1 tháng 9 năm 2008.
- ^ a b Glansdorff, Nicolas; Ying Xu; Labedan, Bernard (ngày 9 tháng 7 năm 2008). "The Last Universal Common Ancestor: emergence, constitution and genetic legacy of an elusive forerunner". Biology Direct. Quyển 3 số 29. tr. 29. doi:10.1186/1745-6150-3-29. PMC 2478661. PMID 18613974.
{{Chú thích tạp chí}}: Quản lý CS1: DOI truy cập mở nhưng không được đánh ký hiệu (liên kết) - ^ a b Brocks, Jochen J.; Logan, Graham A.; Buick, Roger; Summons, Roger E. (ngày 13 tháng 8 năm 1999). "Archean Molecular Fossils and the Early Rise of Eukaryotes". Science. Quyển 285 số 5430. tr. 1033–1036. CiteSeerX 10.1.1.516.9123. doi:10.1126/science.285.5430.1033. PMID 10446042.
- ^ a b c Hedges, S. Blair; Blair, Jaime E.; Venturi, Maria L.; Shoe, Jason L. (ngày 28 tháng 1 năm 2004). "A molecular timescale of eukaryote evolution and the rise of complex multicellular life". BMC Evolutionary Biology. Quyển 4. tr. 2. doi:10.1186/1471-2148-4-2. PMC 341452. PMID 15005799.
{{Chú thích tạp chí}}: Quản lý CS1: DOI truy cập mở nhưng không được đánh ký hiệu (liên kết) - ^ Adl, Sina M.; và đồng nghiệp (tháng 9 năm 2012). "The revised classification of eukaryotes" (PDF). Journal of Eukaryotic Microbiology. Quyển 59 số 5. tr. 429–514. doi:10.1111/j.1550-7408.2012.00644.x. PMC 3483872. PMID 23020233. Bản gốc (PDF) lưu trữ ngày 16 tháng 6 năm 2016. Truy cập ngày 18 tháng 2 năm 2020.
- ^ Burki, F. (2014). "The eukaryotic tree of life from a global phylogenomic perspective". Cold Spring Harbor Perspectives in Biology. Quyển 6 số 5. tr. 1–17. doi:10.1101/cshperspect.a016147. PMC 3996474. PMID 24789819.
- ^ Margulis 1981
- ^ Vellai, Tibor; Vida, Gábor (ngày 7 tháng 8 năm 1999). "The origin of eukaryotes: the difference between prokaryotic and eukaryotic cells". Proceedings of the Royal Society B. Quyển 266 số 1428. tr. 1571–1577. doi:10.1098/rspb.1999.0817. PMC 1690172. PMID 10467746.
- ^ Selosse, Marc-André; Abert, Béatrice; Godelle, Bernard (ngày 1 tháng 3 năm 2001). "Reducing the genome size of organelles favours gene transfer to the nucleus". Trends in Ecology & Evolution. Quyển 16 số 3. tr. 135–141. doi:10.1016/S0169-5347(00)02084-X. PMID 11179577.
- ^ Pisani, Davide; Cotton, James A.; McInerney, James O. (tháng 8 năm 2007). "Supertrees Disentangle the Chimerical Origin of Eukaryotic Genomes". Molecular Biology and Evolution. Quyển 24 số 8. tr. 1752–1760. doi:10.1093/molbev/msm095. PMID 17504772.
- ^ Gray, Michael W.; Burger, Gertraud; Lang, B. Franz (ngày 5 tháng 3 năm 1999). "Mitochondrial Evolution". Science. Quyển 283 số 5407. tr. 1476–1481. Bibcode:1999Sci...283.1476G. doi:10.1126/science.283.5407.1476. PMC 3428767. PMID 10066161.
- ^ Rasmussen, Birger; Fletcher, Ian R.; Brocks, Jochen J.; Kilburn, Matt R. (ngày 23 tháng 10 năm 2008). "Reassessing the first appearance of eukaryotes and cyanobacteria". Nature. Quyển 455 số 7216. tr. 1101–1104. Bibcode:2008Natur.455.1101R. doi:10.1038/nature07381. PMID 18948954.
- ^ Tsu-Ming Han; Runnegar, Bruce (ngày 10 tháng 7 năm 1992). "Megascopic eukaryotic algae from the 2.1-billion-year-old negaunee iron-formation, Michigan". Science. Quyển 257 số 5067. tr. 232–235. Bibcode:1992Sci...257..232H. doi:10.1126/science.1631544. PMID 1631544.
- ^ Javaux, Emmanuelle J.; Knoll, Andrew H.; Walter, Malcolm R. (tháng 7 năm 2004). "TEM evidence for eukaryotic diversity in mid-Proterozoic oceans". Geobiology. Quyển 2 số 3. tr. 121–132. doi:10.1111/j.1472-4677.2004.00027.x.
- ^ Butterfield, Nicholas J. (Winter 2005). "Probable Proterozoic fungi". Paleobiology. Quyển 31 số 1. tr. 165–182. doi:10.1666/0094-8373(2005)031<0165:PPF>2.0.CO;2. Truy cập ngày 30 tháng 1 năm 2015.
{{Chú thích tạp chí}}: Quản lý CS1: năm (liên kết) - ^ a b c d e Neiman, Maurine; Jokela, Jukka (2010). "Sex: Advantage". Encyclopedia of Life Sciences. Encyclopedia of Life Sciences. Hoboken, NJ: John Wiley & Sons. doi:10.1002/9780470015902.a0001716.pub2. ISBN 978-0-470-01617-6.
- ^ Holmes & Jobling 1996
- ^ Christie, Peter J. (tháng 4 năm 2001). "Type IV secretion: intercellular transfer of macromolecules by systems ancestrally related to conjugation machines". Molecular Microbiology. Quyển 40 số 2. tr. 294–305. doi:10.1046/j.1365-2958.2001.02302.x. PMC 3922410. PMID 11309113.
- ^ Michod, Richard E.; Bernstein, Harris; Nedelcu, Aurora M. (tháng 5 năm 2008). "Adaptive value of sex in microbial pathogens" (PDF). Infection, Genetics and Evolution. Quyển 8 số 3. tr. 267–285. doi:10.1016/j.meegid.2008.01.002. PMID 18295550. Bản gốc (PDF) lưu trữ ngày 11 tháng 5 năm 2020. Truy cập ngày 18 tháng 2 năm 2020.
- ^ Bernstein, Harris; Bernstein, Carol (tháng 7 năm 2010). "Evolutionary Origin of Recombination during Meiosis". BioScience. Quyển 60 số 7. tr. 498–505. Bibcode:1985BioSc..35..499W. doi:10.1525/bio.2010.60.7.5.
- ^ Johnsborg, Ola; Eldholm, Vegard; Håvarstein, Leiv Sigve (tháng 12 năm 2007). "Natural genetic transformation: prevalence, mechanisms and function". Research in Microbiology. Quyển 158 số 10. tr. 767–778. doi:10.1016/j.resmic.2007.09.004. PMID 17997281.
- ^ a b Bernstein, Bernstein & Michod 2012, tr. 1–50
- ^ Ramesh, Marilee A.; Malik, Shehre-Banoo; Logsdon, John M., Jr. (ngày 26 tháng 1 năm 2005). "A Phylogenomic Inventory of Meiotic Genes: Evidence for Sex in Giardia and an Early Eukaryotic Origin of Meiosis" (PDF). Current Biology. Quyển 15 số 2. tr. 185–191. Bibcode:1996CBio....6.1213A. doi:10.1016/j.cub.2005.01.003. PMID 15668177. Bản gốc (PDF) lưu trữ ngày 26 tháng 2 năm 2005. Truy cập ngày 22 tháng 12 năm 2008.
{{Chú thích tạp chí}}: Quản lý CS1: nhiều tên: danh sách tác giả (liên kết) - ^ a b Otto, Sarah P.; Gerstein, Aleeza C. (tháng 8 năm 2006). "Why have sex? The population genetics of sex and recombination". Biochemical Society Transactions. Quyển 34 số Pt 4. tr. 519–522. doi:10.1042/BST0340519. PMID 16856849.
- ^ Lin Dong; Shuhai Xiao; Bing Shen; Chuanming Zhou (tháng 1 năm 2008). "Silicified Horodyskia and Palaeopascichnus from upper Ediacaran cherts in South China: tentative phylogenetic interpretation and implications for evolutionary stasis". Journal of the Geological Society. Quyển 165 số 1. tr. 367–378. Bibcode:2008JGSoc.165..367D. doi:10.1144/0016-76492007-074. Truy cập ngày 1 tháng 2 năm 2015.
- ^ Hanley, Kathryn A.; Fisher, Robert N.; Case, Ted J. (tháng 6 năm 1995). "Lower Mite Infestations in an Asexual Gecko Compared With Its Sexual Ancestors". Evolution. Quyển 49 số 3. tr. 418–426. doi:10.2307/2410266. JSTOR 2410266.
- ^ Parker, Matthew A. (tháng 9 năm 1994). "Pathogens and sex in plants". Evolutionary Ecology. Quyển 8 số 5. tr. 560–584. doi:10.1007/bf01238258.
- ^ Bernstein, H., Byerly, H., Hopf, F. and Michod, R. (1984). "Origin of sex". Journal of Theoretical Biology. Quyển 110 số 3. tr. 323–351. doi:10.1016/S0022-5193(84)80178-2. PMID 6209512.
{{Chú thích tạp chí}}: Quản lý CS1: nhiều tên: danh sách tác giả (liên kết) - ^ Birdsell & Wills 2003, tr. 27–137
- ^ Bernstein, Hopf & Michod 1987, tr. 323–370
- ^ Bell, Graham; Mooers, Arne O. (1997). "Size and complexity among multicellular organisms" (PDF). Biological Journal of the Linnean Society. Quyển 60 số 3. tr. 345–363. doi:10.1111/j.1095-8312.1997.tb01500.x. Truy cập ngày 2 tháng 2 năm 2015.
- ^ Kaiser, Dale (tháng 12 năm 2001). "Building a multicellular organism". Annual Review of Genetics. Quyển 35. tr. 103–123. doi:10.1146/annurev.genet.35.102401.090145. PMID 11700279.
- ^ Koschwanez, John H.; Foster, Kevin R.; Murray, Andrew W. (ngày 9 tháng 8 năm 2011). "Sucrose Utilization in Budding Yeast as a Model for the Origin of Undifferentiated Multicellularity". PLOS Biology. Quyển 9 số 8. tr. e1001122. doi:10.1371/journal.pbio.1001122. PMC 3153487. PMID 21857801.
{{Chú thích tạp chí}}: Quản lý CS1: DOI truy cập mở nhưng không được đánh ký hiệu (liên kết) - ^ a b c d Butterfield, Nicholas J. (Summer 2000). "Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes". Paleobiology. Quyển 26 số 3. tr. 386–404. doi:10.1666/0094-8373(2000)026<0386:BPNGNS>2.0.CO;2. Truy cập ngày 1 tháng 2 năm 2015.
- ^ Nakagaki, Toshiyuki; Yamada, Hiroyasu; Tóth, Ágota (ngày 28 tháng 9 năm 2000). "Intelligence: Maze-solving by an amoeboid organism". Nature. Quyển 407 số 6803. tr. 470. Bibcode:2000Natur.407..470N. doi:10.1038/35035159. PMID 11028990.
Thư mục
[sửa | sửa mã nguồn]- Arrhenius, Svante (1980) [Arrhenius paper originally published 1903]. "The Propagation of Life in Space". Trong Goldsmith, Donald (biên tập). The Quest for Extraterrestrial Life: A Book of Readings. Foreword by Sir Fred Hoyle. Mill Valley, CA: University Science Books. Bibcode:1980qel..book...32A. ISBN 978-0-935702-02-6. LCCN 79057423. OCLC 7121102.
- Bengtson, Stefan (2004). "Early Skeletal Fossils" (PDF). Trong Lipps, Jere H.; Waggoner, Benjamin M. (biên tập). Neoproterozoic-Cambrian Biological Revolutions: Presented as a Paleontological Society Short Course at the Annual Meeting of the Geological Society of America, Denver, Colorado, ngày 6 tháng 11 năm 2004. Paleontological Society Papers. Quyển 10. New Haven, CT: Yale University Reprographics & Imaging Service; Paleontological Society. OCLC 57481790. Bản gốc (PDF) lưu trữ ngày 11 tháng 2 năm 2017.
- Bennett, Jeffrey O. (2008). Beyond UFOs: The Search for Extraterrestrial Life and Its Astonishing Implications for Our Future. Princeton, NJ: Princeton University Press. ISBN 978-0-691-13549-6. LCCN 2007037872. OCLC 172521761.
- Benton, Michael J. (1997). Vertebrate Palaeontology (ấn bản thứ 2). Luân Đôn: Chapman & Hall. ISBN 978-0-412-73800-5. OCLC 37378512.
- Benton, Michael J. (2005) [Originally published 2003]. When Life Nearly Died: The Greatest Mass Extinction of All Time (ấn bản thứ 1). Luân Đôn: Thames & Hudson. ISBN 978-0-500-28573-2. LCCN 2002109744. OCLC 62145244.
- Benton, Michael J. (2005a). Vertebrate Palaeontology (ấn bản thứ 3). Malden, MA: Blackwell Science. ISBN 978-0-632-05637-8. LCCN 2003028152. OCLC 53970617.
- Bernstein, Harris; Bernstein, Carol; Michod, Richard E. (2012). "DNA Repair as the Primary Adaptive Function of Sex in Bacteria and Eukaryotes". Trong Kimura, Sakura; Shimizu, Sora (biên tập). DNA Repair: New Research. Hauppauge, NY: Nova Science Publishers. ISBN 978-1-62100-808-8. LCCN 2011038504. OCLC 828424701. Bản gốc lưu trữ ngày 29 tháng 10 năm 2013. Truy cập ngày 7 tháng 11 năm 2018.
- Bernstein, Harris; Hopf, Frederic A.; Michod, Richard E. (1987). "The Molecular Basis of the Evolution of Sex". Trong Scandalios, John G. (biên tập). Molecular Genetics of Development. Advances in Genetics. San Diego, CA: Academic Press. ISBN 978-0-12-017624-3. OCLC 646754753. PMID 3324702.
- Birdsell, John A.; Wills, Christopher (2003). "The Evolutionary Origin and Maintenance of Sexual Recombination: A Review of Contemporary Models". Trong MacIntyre, Ross J.; Clegg, Michael T. (biên tập). Evolutionary Biology. Evolutionary Biology. Quyển 33. New York: Springer Science+Business Media. ISBN 978-1-4419-3385-0. OCLC 751583918.
- Briggs, Derek E. G.; Crowther, Peter R., biên tập (2001). Palaeobiology II. Foreword by E. N. K. Clarkson. Malden, MA: Blackwell Science. ISBN 978-0-632-05149-6. LCCN 0632051477. OCLC 43945263.
- Cairns-Smith, A. G. (1968). "An Approach to a Blueprint for a Primitive Organism". Trong Waddington, C. H (biên tập). Towards a Theoretical Biology. Quyển 1. Edinburgh, Scotland: Edinburgh University Press. ISBN 978-0-85224-018-2. LCCN 71419832. OCLC 230043266.
- Clancy, Paul; Brack, André; Horneck, Gerda (2005). Looking for Life, Searching the Solar System. Cambridge; New York: Cambridge University Press. ISBN 978-0-521-82450-7. LCCN 2006271630. OCLC 57574490.
- Cowen, Richard (2000). History of Life (ấn bản thứ 3). Malden, MA: Blackwell Science. ISBN 978-0-632-04444-3. LCCN 99016542. OCLC 47011068.
- Dalrymple, G. Brent (1991). The Age of the Earth. Stanford, CA: Stanford University Press. ISBN 978-0-8047-1569-0. LCCN 90047051. OCLC 22347190.
- Futuyma, Douglas J. (2005). Evolution. Sunderland, MA: Sinauer Associates. ISBN 978-0-87893-187-3. LCCN 2004029808. OCLC 57311264.
- Gauthier, Jacques; Cannatella, David C.; de Queiroz, Kevin; và đồng nghiệp (1989). "Tetrapod phylogeny". Trong Fernholm, Bo; Bremer, Kåre; Jörnvall, Hans (biên tập). The Hierarchy of Life: Molecules and Morphology in Phylogenetic Analysis. International Congress Series. Quyển 824. Amsterdam, the Netherlands; New York: Excerpta Medica/Elsevier Science Publishers B.V. (Biomedical Division). ISBN 978-0-444-81073-1. LCCN 89001132. OCLC 19129518. "Proceedings from Nobel Symposium 70 held at Alfred Nobel's Björkborn, Karlskoga, Sweden, August 29 – ngày 2 tháng 9 năm 1988"
- Gee, Henry, biên tập (2000). Shaking the Tree: Readings from Nature in the History of Life. Chicago, IL: University of Chicago Press. ISBN 978-0-226-28497-2. LCCN 99049796. OCLC 42476104.
- Gould, Stephen Jay (1989). Wonderful Life: The Burgess Shale and the Nature of History (ấn bản thứ 1). New York: W. W. Norton & Company. ISBN 978-0-393-02705-1. LCCN 88037469. OCLC 18983518.
- Grimaldi, David; Engel, Michael S. (2005). Evolution of the Insects. Cambridge; New York: Cambridge University Press. ISBN 978-0-521-82149-0. LCCN 2004054605. OCLC 56057971.
- Hinde, Rosalind T. (2001). "The Cnidaria and Ctenophora". Trong Anderson, D. T (biên tập). Invertebrate Zoology (ấn bản thứ 2). Melbourne; New York: Oxford University Press. ISBN 978-0-19-551368-4. LCCN 2002276846. OCLC 49663129.
- Holmes, Randall K.; Jobling, Michael G. (1996). "Genetics". Trong Baron, Samuel (biên tập). Medical Microbiology (ấn bản thứ 4). Galveston, TX: University of Texas Medical Branch. Exchange of Genetic Information. ISBN 978-0-9631172-1-2. LCCN 95050499. OCLC 33838234. PMID 21413277.
- Krumbein, Wolfgang E.; Brehm, Ulrike; Gerdes, Gisela; và đồng nghiệp (2003). "Biofilm, Biodictyon, Biomat Microbialites, Oolites, Stromatolites Geophysiology, Global Mechanism, Parahistology" (PDF). Trong Krumbein, Wolfgang E.; Paterson, David M.; Zavarzin, Georgii A. (biên tập). Fossil and Recent Biofilms: A Natural History of Life on Earth. Dordrecht, the Netherlands: Kluwer Academic Publishers. ISBN 978-1-4020-1597-7. LCCN 2003061870. OCLC 52901566. Bản gốc (PDF) lưu trữ ngày 6 tháng 1 năm 2007.
- Labandeira, Conrad C.; Eble, Gunther J. (1999). "The Fossil Record of Insect Diversity and Disparity" (PDF). Trong Anderson, John M.; Thackeray, John Francis; và đồng nghiệp (biên tập). Towards Gondwana Alive: Promoting biodiversity & stemming the Sixth Extinction. Pretoria: Gondwana Alive Society. ISBN 978-1-919795-43-0. LCCN 2001385090. OCLC 44822625. Bản gốc (PDF) lưu trữ ngày 12 tháng 4 năm 2016. Truy cập ngày 7 tháng 11 năm 2018. "Preview booklet for 'Gondwana alive: biodiversity and the evolving terrestrial biosphere', book planned for September 2000, and associated projects."
- Leakey, Richard (1994). The Origin of Humankind. Science Masters Series. New York: Basic Books. ISBN 978-0-465-03135-1. LCCN 94003617. OCLC 30739453.
- Margulis, Lynn (1981). Symbiosis in Cell Evolution: Life and its Environment on the Early Earth. San Francisco, CA: W. H. Freeman and Company. ISBN 978-0-7167-1256-5. LCCN 80026695. OCLC 6982472.
- McKinney, Michael L. (1997). "How do rare species avoid extinction? A paleontological view". Trong Kunin, William E.; Gaston, Kevin J. (biên tập). The Biology of Rarity: Causes and consequences of rare—common differences (ấn bản thứ 1). Luân Đôn; New York: Chapman & Hall. ISBN 978-0-412-63380-5. LCCN 96071014. OCLC 36442106.
- Miller, G. Tyler; Spoolman, Scott E. (2012). Environmental Science (ấn bản thứ 14). Belmont, CA: Brooks/Cole. ISBN 978-1-111-98893-7. LCCN 2011934330. OCLC 741539226.
- Newman, William L. (ngày 9 tháng 7 năm 2007). "Age of the Earth". Geologic Time. Reston, VA: Publications Services, USGS. OCLC 18792528. Truy cập ngày 29 tháng 8 năm 2008.
- O'Leary, Margaret R. (2008). Anaxagoras and the Origin of Panspermia Theory. Bloomington, IN: iUniverse. ISBN 978-0-595-49596-2. OCLC 757322661.
- Padian, Kevin (2004). "Basal Avialae". Trong Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (biên tập). The Dinosauria (ấn bản thứ 2). Berkeley: University of California Press. ISBN 978-0-520-24209-8. LCCN 2004049804. OCLC 55000644.
- Sansom, Ivan J.; Smith, Moya M.; Smith, M. Paul (2001). "The Ordovician radiation of vertebrates". Trong Ahlberg, Per Erik (biên tập). Major Events in Early Vertebrate Evolution: Palaeontology, phylogeny, genetics and development. Systematics Association special volume series. Quyển 61. Luân Đôn; New York: Taylor & Francis. ISBN 978-0-415-23370-5. LCCN 00062919. OCLC 51667292.
- Thewissen, J. G. M.; Madar, S. I.; Hussain, S. T. (1996). Ambulocetus natans, an Eocene cetacean (Mammalia) from Pakistan. Courier Forschungsinstitut Senckenberg. Quyển 191. Frankfurt: Senckenbergische Naturforschende Gesellschaft. ISBN 978-3-929907-32-2. LCCN 97151576. OCLC 36463214.
Đọc thêm
[sửa | sửa mã nguồn]- Dawkins, Richard (1989). The Selfish Gene . Oxford; New York: Oxford University Press. ISBN 978-0-19-286092-7. LCCN 89016077. OCLC 20012195.
- Dawkins, Richard (2004). The Ancestor's Tale: A Pilgrimage to the Dawn of Life. Boston: Houghton Mifflin Company. ISBN 978-0-618-00583-3. LCCN 2004059864. OCLC 56617123.
- Ruse, Michael; Travis, Joseph, biên tập (2009). Evolution: The First Four Billion Years. Foreword by Edward O. Wilson. Cambridge, MA: Belknap Press of Harvard University Press. ISBN 978-0-674-03175-3. LCCN 2008030270. OCLC 225874308.
- Smith, John Maynard; Szathmáry, Eörs (1997) [Originally published 1995; Oxford: W. H. Freeman/Spektrum]. The Major Transitions in Evolution. Oxford; New York: Oxford University Press. ISBN 978-0-19-850294-4. LCCN 94026965. OCLC 715217397.
Thông tin chung
[sửa | sửa mã nguồn]- "Evolution". The Virtual Fossil Museum. Truy cập ngày 22 tháng 2 năm 2015. General information on evolution compiled by Roger Perkins
- "Understanding Evolution: your one-stop resource for information on evolution". University of California, Berkeley. Truy cập ngày 22 tháng 2 năm 2015.
- "Evolution Resources". Washington, D.C.: National Academies. Truy cập ngày 23 tháng 2 năm 2015.
- "Tree of Life". Bản gốc lưu trữ ngày 10 tháng 2 năm 2015. Truy cập ngày 23 tháng 2 năm 2015. Tree of life diagram by Neal Olander
- "Evolution". New Scientist. Truy cập ngày 23 tháng 2 năm 2015.
- Brain, Marshall. How Evolution Works tại HowStuffWorks
- "Modern Theories of Evolution: An Introduction to the Concepts and Theories That Led to Our Current Understanding of Evolution". Palomar College. Bản gốc lưu trữ ngày 6 tháng 8 năm 2009. Truy cập ngày 23 tháng 2 năm 2015. Tutorial created by Dennis O'Neil
Lịch sử các quan điểm tiến hóa
[sửa | sửa mã nguồn]- van Wyhe, John (biên tập). "The Complete Work of Charles Darwin Online". Truy cập ngày 23 tháng 2 năm 2015.
- Price, R. G. "Understanding Evolution: History, Theory, Evidence, and Implications". rationalrevolution.net. Truy cập ngày 23 tháng 2 năm 2015.







![[Review Sách] Sống thực tế giữa đời thực dụng - Khi nỗ lực trở thành bản năng](https://down-tx-vn.img.susercontent.com/sg-11134201-7qve1-lj0r8adyezkf95.webp) GIẢM
10%
GIẢM
10%
 GIẢM
20%
GIẢM
20%
 GIẢM
9%
GIẢM
9%
![[Tập hiểu sâu]: Vì sao Bạn luôn KHÔNG ỔN?](https://images.spiderum.com/sp-images/f5ab2630336211eea9e7ef608f381f46.png) GIẢM
12%
GIẢM
12%
 GIẢM
25%
GIẢM
25%
 GIẢM
40%
GIẢM
40%


![Một số thông tin về Đại quỷ tộc [Ogre] (Quỷ lớn) Tensura](https://1.bp.blogspot.com/-35QVuGjxuqo/XojJflBytoI/AAAAAAAAAP8/XtXaH2bNZ5Q5p5cSyW4bZRidK6ku386UgCLcBGAsYHQ/w700-h408-p-k-no-nu/tensuraova.jpg)
