Nấm
| Nấm | |
|---|---|
| Thời điểm hóa thạch: Đầu kỷ Devon - gần đây (xem thêm trong văn bản) | |
 Theo chiều kim đồng hồ từ góc trên bên trái: Nấm diệt ruồi Amanita muscaria (Basidiomycota-nấm đảm); Sarcoscypha coccinea (Ascomycota-nấm nang); mốc đen (Zygomycota-nấm tiếp hợp); nấm roi/ nấm trứng(nấm rơm) (Chytridiomycota); Penicillium conidiophore. | |
| Phân loại khoa học | |
| Liên vực (superdomain) | Neomura |
| Vực (domain) | Eukaryota |
| Nhánh | Unikonta |
| Nhánh | Opisthokonta |
| Giới (regnum) | Fungi (L., 1753) R.T. Moore, 1980[1] |
| Phân giới/Ngành/Phân ngành[2] | |
| |
Giới Nấm (tên khoa học: Fungi) bao gồm những sinh vật nhân chuẩn dị dưỡng có thành tế bào bằng kitin (chitin). Phần lớn nấm phát triển dưới dạng các sợi đa bào được gọi là sợi nấm (hyphae) tạo nên thể sợi (mycelium), một số nấm khác lại phát triển dưới dạng đơn bào. Quá trình sinh sản (hữu tính hoặc vô tính) của nấm thường là qua bào tử, được tạo ra trên những cấu trúc đặc biệt hay thể quả. Một số loài mất khả năng tạo nên những cấu trúc sinh sản đặc biệt và nhân lên qua hình thức sinh sản sinh dưỡng.
Những đại diện tiêu biểu của nấm là nấm mốc, nấm men và nấm lớn (nấm thể quả). Giới Nấm là nhóm sinh vật đơn ngành (monophyletic) mà có nguồn gốc hoàn toàn khác biệt với những sinh vật có hình thái tương tự như nấm nhầy (myxomycetes) hay mốc nước (oomycetes). Nấm có mối quan hệ gần với động vật hơn là thực vật, cho dù thế thì môn học về nấm, hay nấm học, lại thường được xếp vào thành một nhánh của môn thực vật học.
Trên Trái Đất, đa phần các nấm đều không thể nhìn thấy được bằng mắt thường, chúng sống phần lớn ở trong đất, chất mùn, xác sinh vật chết, cộng sinh hoặc ký sinh trên cơ thể động, thực vật và nấm khác. Vi nấm đóng 1 vai trò quan trọng trong hệ sinh thái, chúng phân hủy các vật chất hữu cơ và không thể thiếu được trong chu trình chuyển hóa và trao đổi vật chất. Một số loài nấm có thể nhận thấy được khi ở dạng quả thể, như nấm lớn và nấm mốc. Nấm được ứng dụng rất rộng rãi trong đời sống lẫn sản xuất. Nhiều loài được sử dụng trong công nghệ thực phẩm, sử dụng làm thức ăn hoặc trong quá trình lên men. Nấm còn được dùng để sản xuất chất kháng sinh, hoóc môn trong y học và nhiều loại enzym, ngày nay nhiều loại nấm được biết đến và sử dụng trong phòng chống nhiều loại bệnh hiểm nghèo như viêm gan, mỡ máu, đột quỵ, ung thư... Tuy vậy, nhiều loại nấm lại có chứa các chất hoạt động sinh học được gọi là mycotoxin, như ancaloit và polyketit-là những chất độc đối với động vật lẫn con người (gây buồn nôn, khó chịu, ngộ độc). Một số loại nấm được sử dụng để kích thích hoặc dùng trong các nghi lễ truyền thống với vai trò tác động lên trí tuệ và hành vi của con người. Vài loại nấm có thể gây ra các chứng bệnh cho con người và động vật, cũng như bệnh dịch cho cây trồng, mùa màng và có thể gây tác động lớn lên an ninh lương thực và kinh tế.
Sự đa dạng
[sửa | sửa mã nguồn]Nấm phân bố trên toàn thế giới và phát triển ở nhiều dạng môi trường sống khác nhau, kể cả sa mạc, nơi tập trung nồng độ muối cao[3] hay có bức xạ ion hóa,[4] cũng như trầm tích biển sâu.[5] Đa phần nấm sống ở trên cạn, nhưng một số loài sống ở môi trường nước (như Batrachochytrium dendrobatidis - ký sinh và làm suy giảm số lượng động vật lưỡng cư toàn cầu). Nấm thủy sinh còn sống ở vùng nhiệt dịch đại dương.[6] Nấm và vi khuẩn là những sinh vật phân giải chính có vai trò quan trọng đối với các hệ sinh thái trên cạn trên toàn thế giới. Dựa theo sự theo tỉ lệ giữa số loài nấm với số loài thực vật ở trong cùng một môi trường, người ta ước tính giới Nấm có khoảng 1,5 triệu loài[7]. Khoảng 100.000 loài nấm đã được các nhà phân loại học phát hiện và miêu tả,[8] tuy nhiên kích cỡ thực sự của tính đa dạng của giới Nấm vẫn còn là điều bí ẩn[9]. Đa phần nấm phát triển dưới dạng các sợi đa bào gọi là sợi nấm, cấu tạo nên thể sợi (hay khuẩn ty), trong khi những loài khác thì lại phát triển dưới dạng đơn bào[10][11]. Cho đến gần đây, nhiều loại nấm đã được miêu tả dựa trên những đặc điểm hình thái, như kích cỡ và hình dạng các bào tử hay quả thể, hay dựa trên khái niệm loài sinh vật với sự trợ giúp của các công cụ phân tử, như phương pháp Dideoxy, đã gia tăng mạnh cách thức và khả năng ước tính sự đa dạng của nấm trong phạm vi các nhóm phân loại khác nhau[12].
Sinh thái
[sửa | sửa mã nguồn]
Dù không dễ thấy, nhưng nấm lại có mặt ở tất cả các môi trường trên Trái Đất và đóng một vai trò rất quan trọng trong hệ sinh thái. Cùng với vi khuẩn, nấm là sinh vật phân hủy chính ở hầu hết các hệ sinh thái trên cạn (và một số là ở dưới nước), bởi vậy nên chúng cũng có vai trò quan trọng các chu trình sinh địa hóa và ở nhiều lưới thức ăn. Khi sống hoại sinh hay cộng sinh, chúng phân hủy những vật chất hữu cơ thành những phân tử vô cơ, rồi sau đó những chất này sẽ được đồng hóa ở thực vật hay những sinh vật khác.[13][14]
Cộng sinh
[sửa | sửa mã nguồn]Nấm có mối quan hệ cộng sinh với hầu hết tất cả các giới [15][16][17]. Quan hệ của chúng có thể hỗ trợ hoặc đối nghịch nhau, hay với những nấm hội sinh thì không đem lại bất cứ lợi ích hay tác hại rõ ràng nào đối với vật chủ.[18][19][20]
Với thực vật
[sửa | sửa mã nguồn]
Nấm rễ là một hình thức cộng sinh giữa thực vật và nấm, chia làm hai loại: nấm rễ trong (endomycorrhiza, tức nấm ký sinh đơn bào sống bên trong tế bào rễ cây) và nấm rễ ngoài (ectomycorrhiza, tức rễ của nấm bám dày đặc xung quanh đầu rễ cây và xâm nhập vào giữa các tế bào rễ cây). Đây là quần hợp nấm-thực vật được biết nhiều nhất và đóng vai trò quan trọng trong quá trình phát triển của thực vật cũng như nhiều hệ sinh thái, hơn 90% các loài thực vật có quan hệ với nấm theo hình thức nấm rễ và phụ thuộc vào mối quan hệ này để tồn tại[21][22][23]. Sự cộng sinh nấm rễ đã có lịch sử xa xưa, ít nhất là từ hơn 400 triệu năm về trước [24]. Chúng thường làm tăng khả năng hấp thu các hợp chất vô cơ của thực vật, như nitrat và phosphat, từ những đất có nồng độ những nguyên tố thiết yếu thấp[25]. Ở một số nấm rễ, thành phần nấm có thể đóng vai trò trung gian giữa thực vật với thực vật, vận chuyển carbohydrate và các chất dinh dưỡng khác. Những cộng đồng nấm rễ đó được gọi là "mạng lưới nấm rễ chung"[26]. Một số nấm có khả năng kích thích sự sinh trưởng của cây bằng cách tiết ra các hoóc môn thực vật như axít idolaxetic (IAA) [27].
Địa y là dạng cộng sinh giữa nấm (hầu hết các loài nấm nang và một số nấm đảm) với tảo hay vi khuẩn lam (gọi chung là đối tác quang hợp), trong đó những tế bào quang hợp được gắn vào những mô nấm [28]. Giống với nấm rễ, những đối tác quang hợp sẽ cung cấp cacbohyđrat được tạo ra trong quá trình quang hợp, đổi lại nấm cung cấp cho chúng các chất khoáng và nước. Những chức năng của toàn bộ cơ thể địa y gần như giống hệt với một cơ thể đơn độc. Địa y là những sinh vật tiên phong và xuất hiện ở những nơi nguyên thủy như đá tảng hay nham thạch núi lửa đã nguội. Chúng có thể thích nghi cực tốt với những điều kiện khắc nghiệt như giá lạnh hay khô hạn và là những ví dụ tiêu biểu nhất của sự cộng sinh[27].
Một số loài nấm sống trong cây có thể tiết ra những độc tố nấm để ngăn cản những động vật ăn cỏ ăn vật chủ của chúng[29].
Với côn trùng
[sửa | sửa mã nguồn]Nhiều côn trùng có mối quan hệ hỗ trợ với nhiều loại nấm. Vài loại kiến trồng những loài nấm thuộc bộ Nấm mỡ (Agaricales) để làm nguồn thức ăn chính, trong khi đó những loài bọ cánh cứng Ambrosia trồng nhiều loài nấm trong lớp vỏ cây mà chúng cư trú [30]. Loài mối ở xavan châu Phi cũng được biết có khả năng trồng nấm [31].
Mầm bệnh và ký sinh
[sửa | sửa mã nguồn]
Tuy vậy, nhiều loại nấm lại ký sinh trên con người, thực vật, động vật và nấm khác. Những loài nấm gây bệnh trên cây trồng có thể gây thiệt hại rộng lớn cho ngành nông nghiệp và lâm nghiệp, ví dụ như nấm đạo ôn (Magnaporthe oryzae) gây bệnh cho lúa[32], Ophiostoma ulmi và Ophiostoma novo-ulmi gây ra bệnh du Hà Lan[33], còn Cryphonectria parasitica là nguyên nhân của bệnh thối cây dẻ[34]. Những loài gây bệnh cho cây thuộc các chi Fusarium, Ustilago, Alternaria và Cochliobolus[19], còn những loài có khả năng gây bệnh cho người lại thuộc các chi như Aspergillus, Candida, Cryptoccocus[20][35][36], Histoplasma[37] và Pneumocystis [38]. Chúng có thể gây ra những bệnh ngoài da ở người như nấm chân hay hắc lào cho đến những bệnh nguy hiểm có thể gây chết người như viêm màng não (nấm Cryptococcus neoformans)[39] hay viêm phổi. Nấm gây ra nhiều bệnh cơ hội, tức những bệnh tấn công những người bị suy giảm miễn dịch [40], trong đó có những người bị HIV/AIDS [41], ví dụ như bệnh candidiasis (nấm Candida, gây ra chứng lở miệng ở trẻ em và âm đạo phụ nữ), histoplasmosis (Histoplasma capsulatum), cryptococcosis (Cryptococcus neoformans), aspergillosis (Aspergillus), coccidioidomycosis (Coccidioides immitis hay C. posadasii), viêm phổi pneumocystis (Pneumocystis jirovecii)... và rất nhiều bệnh khác[41][42].
Có khoảng 70 loài[43] nấm sinh bào tử là những tác nhân gây dị ứng. Chúng có thể là nấm mốc trong nhà hay ngoài trời, đa phần là nấm sợi như các chi Alternaria, Asperillus, Cladosporium, Helminthosporium, Epicoccum, Penicillium, Fusarium..., chỉ có vài loài là nấm đơn bào như Candida, Rhodotorula, có một số loài là nấm lớn như Agaricus, Coprinus, Fomes, Ganoderma...[43][44] Bào tử nấm có thể gây ra những chứng như hen suyễn, viêm mũi dị ứng[45], các bệnh nấm dị ứng phế quản phổi và viêm phổi quá mẫn[44].
Săn mồi
[sửa | sửa mã nguồn]Một số loài nấm là những kẻ săn giun tròn. Chúng có thể biến đổi sợi nấm để tạo thành những cấu trúc đặc biệt có chức năng bẫy giun tròn, nên được gọi với tên chung là nấm bẫy mồi[46]. Những loại bẫy thường thấy là: mạng dính (hay lưới dính), bọng dính, vòng không thắt, cột dính, vòng thắt[47][48] và bào tử dính[49]. Các loài nấm bắt mồi theo kiểu này thường thuộc các chi Arthrobotrys, Dactylaria, Dactylella và Trichothecium[46][50]. Có vài loài như Zoopage phanera[46][51] thì lại tiết chất dính ra toàn bộ mặt ngoài sợi nấm và cũng có khả năng bẫy mồi tương tự.
Dinh dưỡng và khả năng tự dưỡng
[sửa | sửa mã nguồn]Sự phát triển của nấm dưới dạng sợi nấm ở những môi trường rắn cũng như dưới dạng đơn bào ở môi trường nước, đều được điều chỉnh để hút các chất dinh dưỡng hiệu quả nhất từ môi trường, bởi chúng đều có tỉ lệ diện tích bề mặt trên thể tích cao. Sự thích nghi hình thái đã được bổ sung bởi những enzym thủy phân trong những môi trường tiêu hóa có phân tử hữu cơ lớn, như polysaccarit, protein, lipid và những chất nền dinh dưỡng khác. Những phân tử này bị thủy phân thành những phân tử nhỏ hơn[52][53][54], sau đó trở thành những chất dinh dưỡng được hấp thụ vào tế bào nấm.
Thông thường nấm được coi là những sinh vật dị dưỡng, tức những cơ thể chỉ có thể lấy cacbon từ những sinh vật khác cho quá trình trao đổi chất. Tuy nhiên, nấm đã tiến hóa khả năng chuyển hoá mà cho phép chúng sử dụng đa dạng những loại chất nền hữu cơ để phát triển, bao gồm các hợp chất đơn giản như nitrat, amonia, acetat hay ethanol[55][56]. Những nghiên cứu gần đây cho thấy khả năng rằng nấm đã sử dụng sắc tố melanin để lấy năng lượng từ những phóng xạ ion hóa, như tia gamma, gọi nôm na là "vô tuyến dưỡng"[57]. Người ta cho rằng quá trình này có điểm tương đồng với quá trình quang hợp ở thực vật[57]. Tuy nhiên, hiện nay đang thiếu những bằng chứng sinh hóa có giá trị ủng hộ cho giả thuyết này.
Vai trò đối với con người
[sửa | sửa mã nguồn]Chế biến thực phẩm
[sửa | sửa mã nguồn]

Nấm men bánh mì với loài chính là Saccharomyces cerevisiae, một nấm đơn bào, được sử dụng rộng rãi trong việc làm bánh mì và những sản phấm từ bột mì khác, như pizza hay bánh bao [58]. Trong quá trình lên men rượu dưới điều kiện yếm khí, nấm men sẽ sản sinh ra rượu etanol và khí cacbonic (CO2) có vai trò làm nở bột mì. Một số loài nấm men của chi Saccharomyces cũng được sử dụng để sản xuất đồ uống có cồn, như bia hay rượu vang thông qua quá trình lên men rượu[59]. Nấm men và vi khuẩn acetic được sử dụng trong quá trình chuẩn bị Kombucha, một loại trà ngọt lên men. Những loại men tìm thấy trong trà rất đa dạng, như Brettanomyces bruxellensis, Candida stellata, Schizosaccharomyces pombe, Torulaspora delbrueckii và Zygosaccharomyces bailii[60]. Ngoài ra, Vegemite và Marmite, những chiết xuất nấm men[61], đã được sử dụng rộng rãi làm gia vị để chế biến thức ăn trong đời sống.

Một số loài nấm sợi thường được sử dụng để sản xuất sinh khối protein. Mốc Aspergillus oryzae được sử dụng rộng rãi ở nhiều nước, như sản xuất nước tương (xì dầu), súp miso và rượu sake ở Nhật Bản[62] hay làm tempeh ở Java[63]. Mốc hoa cau được dùng để sản xuất tương, loại thực phẩm phổ biến ở Việt Nam, cũng chính là A. oryzae[62], tuy nhiên tương sản xuất thủ công lại có độ an toàn không cao, bởi những loại mốc tốt và không độc như A. oryzae và Aspergillus sojae lại rất dễ lẫn lộn với những loại mốc nguy hiểm có độc tố gây ung thư khác là A. flavus và A. parasiticus[62]. Quorn là loại thực phẩm giàu protein được sản xuất từ mốc Fusarium venenatum[64], và được dùng trong việc chế biến đồ ăn chay. Nhiều loại thực phẩm khác cũng được chế biến bởi mốc như chao (ủ nhờ mốc Actinomucor elegans, Mucor racemosus hay Rhizopus)[65], ang-kak (gạo lên men với mốc đỏ Monascus purpureus)[66], salami (một loại xúc xích, lên men nhờ P. nalgiovense, P. chrysogenum)[67][68]. Trong sản xuất pho mát, một kinh nghiệm thông thường là cấy bào tử nấm vào sữa đông để tạo mốc, việc này sẽ cho ra hương vị và kết cấu đặc biệt độc nhất của pho mát. Ở những loại pho mát xanh, như Stilton hay Roquefort, thì những vân xanh được tạo ra bởi loài nấm Penicillium roqueforti[69].
Nấm ăn và nấm độc
[sửa | sửa mã nguồn]

Những loài nấm quả thể được biết đến với hai dạng: nấm ăn được và nấm độc. Nấm ăn được sử dụng rộng rãi làm thực phẩm, chúng có thể sử dụng trong rất nhiều món ăn, ở nhiều nền ẩm thực khác nhau. Nấm là thực phẩm có giá trị dinh dưỡng cao, có độ đạm cao và ít chất béo, chứa nhiều vitamin nhóm B và C[65]. Dù nấm không phải là nguốn vitamin D đáng kể, nhưng hàm lượng vitamin D có thể tăng lên khi được phơi với ánh sáng (nhất là tia cực tím) dù điều này làm thẫm lớp vỏ của chúng[70]. Nấm cũng chứa nhiều nguyên tố vi lượng, như sắt, seleni, natri, kali, magnesi và phosphor[65].
Những loại nấm ăn được thường xuyên bày bán ở các chợ và siêu thị đều được trồng ở các trang trại nấm. Loại nấm phổ biến nhất là nấm mỡ (Agaricus bisporus)[71], được trồng ở ít nhất 70 quốc gia trên thế giới[72]. Những dạng khác của A.bisporus là portabella và nấm mũ (crimini) cũng được trồng thương mại. Nhiều loại nấm châu Á cũng được trồng và tiêu thụ rộng rãi là nấm rơm (Volvariella volvacea), nấm hương (Lentinula edodes), nấm sò (Pleurotus ostreatus), mộc nhĩ (Auricularia auricula-judae), enokitake (nấm kim châm, Flammulina) và nấm múa (Grifola frondosa).

Có nhiều loại nấm được thu hoạch từ tự nhiên để cho cá nhân hay để bán như nấm sữa (Lactarius deliciosus), nấm nhăn (nấm bụng dê, Morchella), nấm mồng gà (Cantharellus), nấm cục (Tuber), nấm kèn đồng (Cantharellus) và nấm thông (Boletus edulis), chúng thường đắt tiền và dành cho những người sành ăn. Hái nấm là hoạt động phổ biến ở nhiều vùng của Châu Âu và tây bắc Hoa Kỳ. Tuy nhiên những người đi hái nấm phải rất chú trọng về việc phân biệt nấm ăn và nấm độc. Có nhiều loại nấm đặc biệt độc đối với con người, độc tính của nấm có thể nhẹ và gây ra bệnh tiêu hóa hay dị ứng cũng như ảo giác, nhưng cũng có thể đủ mạnh để gây liệt các cơ quan và chết người. Có khoảng 10.000 loại nấm thịt, trong đó có một nửa là ăn được và 100 loài có độc tố cao[65]. Những loại nấm gây chết người thuộc về các chi Inocybe, Entoloma, Hebetoma, Cortinarius và nổi tiếng nhất là Amanita[73]. Những loài thuộc chi cuối như "thiên thần hủy diệt" A. virosa hay nấm tử thần A. phalloides là những loại nấm độc chết người thông dụng nhất[74]. Loại nấm moscela giả (Gyromitra esculenta) khi nấu chín là một thức ăn ngon, nhưng lại độc khi ăn sống[75]. Nấm Tricholoma equestre đã từng được cho là ăn được cho đến khi nó bị phát hiện là gây ra bệnh Rhabdomyolysis (hủy hoại cơ bắp)[76].

Nấm màu đỏ Amanita muscaria gây độc không thường xuyên, khi ăn vào nó có thể trở thành loại thuốc kích thích và sinh ảo giác. Trong lịch sử, những tu sĩ cổ đại người Celt ở Bắc Âu và người Koryak ở Siberi đã sử dụng loại nấm này với mục đích tôn giáo và làm phép[77]. Cũng có nhiều loài nấm gây ảo giác khác, chúng được gọi là "nấm ma thuật", "mush" hoặc "shroom", thuộc nhiều chi khác nhau như Psilocybe, Panaeolus, Gymnopilus, Copelandia, Conocybe... Chúng có thể tác động lên trí tuệ và hành vi của con người, tạo cảm giác hư ảo hưng phấn, và cũng có vai trò trong việc chữa trị truyền thống ở một số địa phương.
Để phân biệt được nấm ăn được và nấm độc thì có nhiều kinh nghiệm cùng với các cơ sở nghiên cứu: không ăn nấm sặc sỡ, có mùi hắc, không ăn nấm quá non hay quá già, không ăn nấm có chảy sữa...[78] Tuy nhiên có rất nhiều trường hợp ngoại lệ, như có nấm độc có màu sắc và hình dạng giống nấm thường[79]. Vì thế rất khó để có thể nhận biết được nấm an toàn và nấm độc nếu không có đủ kinh nghiệm và kiến thức, và lời khuyên tốt nhất là hãy coi tất cả nấm ở trong rừng là nấm độc và bỏ qua nó.
Dược liệu và chiết xuất
[sửa | sửa mã nguồn]Nhiều loại nấm ăn đã được sử dụng trong y học truyền thống hàng ngàn năm nay. Những loại nấm như nấm múa, nấm hương (đông cô), nấm chaga, nấm linh chi... đã được tập trung nghiên cứu bởi khả năng chống ung thư, chống virus và tăng cường hệ miễn dịch của chúng. Loài nấm Đông trùng hạ thảo (Cordyceps sinensis) được coi là một dược liệu quý hiếm và đã được sử dụng ở Trung Quốc từ lâu [80]. Loài nấm cổ linh chi (Ganoderma applanatum) cũng từng được coi là một "thần dược" ở Việt Nam, mặc dù không có bằng chứng cụ thể nào về khả năng trị bệnh của nó [81]. Psilocybin và LSD, những chất gây ảo giác được chiết xuất từ nấm, có thể dùng để chữa các bệnh về tâm thần, như chứng rối loạn ám ảnh cưỡng chế (OCD)[82][83], và cũng được dùng (với lượng nhỏ) để chấp dứt những cơn nhức đầu hàng loạt (cluster headache) hay đau nửa đầu[84]. LSD mạnh gấp 100 lần psilocybin [85], được tổng hợp vào năm 1938, và là một loại ma túy quen thuộc.

Trong số 12.000 loại kháng sinh được biết năm 1995 có khoảng 22% được sản xuất từ nấm sợi[86]. Trong số đó, kháng sinh Penicillin, được Alexander Fleming tổng hợp từ nấm Penicillium chrysogenum vào năm 1928, được sử dụng rất rộng rãi trong chữa trị y học thế kỷ XX. Chúng có thể chữa được các bệnh vi khuẩn như bạch hầu, viêm phổi, viêm màng não, hôi miệng, giang mai, lậu và kể cả vi khuẩn Staphylococcus gây ra nhiễm trùng huyết [27]. Một loại kháng sinh β-lactam phổ biến khác, Cephalosporin, cũng được tổng hợp năm 1948 từ nấm Cephalosporium acremonium[87].
Áp dụng công nghệ DNA tái tổ hợp, nhiều gen đã được chuyển vào những loại nấm như nấm men S. cerevisiae, Pichia pastoris, Schizosaccharomyces pombe[88], Kluyveromyces lactis, Candida albicans, Hansenula polymorpha, Yarrowia lipolytica.. và nấm sợi Aspergillus niger, A. nidulans, A. oryzae, Neurospora crassa, Trichoderma reesei...[89] với mục đích sản xuất công nghiệp. Nhờ khả năng phát triển nhanh, chúng được nuôi trồng và cho ra nhiều loại sản phẩm protein đa dạng giá trị rất lớn trong y học như insulin[88][90], vắc-xin viêm gan B, interferon, nhân tố tăng trưởng biểu bì, hemoglobin người, superoxide dismutase, interleukin[91]... Ngoài ra các nấm chuyển gen cùng với các vi khuẩn còn những nguồn chính cho các enzym sử dụng trong công nghệ thực phẩm[91].
Việc nuôi cấy nấm men trong sản xuất công nghiệp ngoài những sản phẩm có được do chuyển gen, còn có rất nhiều sản phẩm khác sử dụng trong thực phẩm, dinh dưỡng và y tế như cồn, các acid hữu cơ (acid citric, acetic, lactic, malic, gluconic, fumaric, itaconic..), vitamin (β-carotene, riboflavin, acid béo thiết yếu), enzym (protease, amylase, glucoamylase, men dịch vị..), pullulan, các este, polysaccharide, lipid, glycolipid, các hoạt chất thứ cấp như kháng sinh, chất ức chế miễn dịch (Ciclosporin A từ Tolypocladium inflatum), chất làm giảm cholestrol (lovastatin, pravastatin), chất chống ung thư (Taxol), chất màu... và nhiều chất hóa học khác[91]. Thậm chí những cellulase và xylanase mà nấm tiết ra còn được sử dụng trong sản xuất quần jeans stone-washed[92] cũng như trong công nghiệp sản xuất giấy và bột giấy[93]. Ngành công nghiệp nuôi cấy nấm đã trở thành một bộ phận quan trọng trong nền công nghiệp toàn cầu[91].
Phục hồi sinh học
[sửa | sửa mã nguồn]Thuật ngữ phục hồi sinh học bằng nấm (mycorestoration) đã được nhà nấm học Paul Stamets sử dụng trong cuốn sách Mycelium Running: How Mushrooms Can Help Save the World (Ten Speed Press, 2005) [94], trong đó ông nêu 4 dạng: lọc sinh học bằng nấm (mycofiltration), lâm nghiệp bằng nấm (mycoforestry), cải tạo sinh học bằng nấm (mycoremediation) và thuốc trừ sâu bằng nấm (mycopesticide)[95]. Về nấm được dùng làm thuốc trừ sâu sinh học, có thể xem ở điều khiển sinh học bằng nấm.
Cải tạo sinh học được hiểu là sử dụng các vi sinh vật, nấm hay cây xanh để đưa môi trường tự nhiên bị ô nhiễm trở về trạng thái ban đầu. Nấm tiết ra bên ngoài các enzym và axít có khả năng phân giải lignin và cellulose, hai thành phần chính của gỗ, và có cấu trúc tương tự với nhiều chất ô nhiễm hữu cơ. Một số loại nấm, đặc biệt là nấm mục trắng (Phanerochaete chrysosporium), có thể phân giải các chất độc hại cho môi trường như thuốc trừ sâu, thuốc diệt cỏ, pentachlorophenol, creosote, nhựa than đá và những nhiên liệu nặng thành CO2, nước và những nguyên tố cơ bản [96]. Chúng có thể phân giải hay loại trừ các nguyên tố độc hại như chì, arsenic, caesi, cadmi, thủy ngân, những vũ khí và chất độc hóa học như thuốc nổ TNT, chất độc thần kinh VX và sarin[97]. Những nghiên cứu gần đây cho thấy nấm có khả năng bao bọc urani vào trong những dạng khoáng[98].
Điều khiển sinh học
[sửa | sửa mã nguồn]
Nấm rất tích cực trong cạnh tranh về dinh dưỡng và không gian với những sinh vật khác, thông qua định luật Gause (nguyên tắc ức chế cạnh tranh)[99], và điều này đôi khi có lợi cho con người. Ví dụ, nấm có thể ngăn chặn sự tăng trưởng hay loại trừ kẻ thù nguy hiểm của thực vật và con người, như kiến đục gỗ, mối, châu chấu, muỗi, ve bét [100], cỏ dại, giun tròn hay nấm khác mà có thể gây hại cho mùa màng và nhà cửa [101]. Khả năng điều khiển sinh học các loài gây hại cho nông nghiệp của nấm đã được quan tâm và ứng dụng thực tế. Loài nấm ký sinh côn trùng đã được sử dụng làm thuốc trừ sâu sinh học vì khả năng ký sinh và tiêu diệt côn trùng của chúng [102]. Một số ví dụ về những nấm loại này là Beauveria bassiana, Metarhizium anisopliae, chi Hirsutella, chi Paecilomyces và Verticillium lecanii[103][104]. Có ít nhất 14 loại nấm có khả năng chống rệp [105]. Loài nấm thuộc chi Trichoderma cũng có khả năng ngăn chặn những loài nấm gây bệnh cho cây [106].
Những loài nấm sống trong cây thuộc chi Neotyphodium như N. coenophialum tiết ra các ancaloit có độc tính với các loài động vật ăn thực vật có xương sống hay không xương sống, mặc dù một số chất có thể độc với những loài gia súc như cừu [107]. Đặc tính của các loại nấm này đã được sử dụng trong các chương trình gây giống cây, nhằm tăng khả năng chống chịu và phòng ngự của cây trồng với các loài ăn thực vật[108].
Kẻ phá hoại
[sửa | sửa mã nguồn]
Mặc dù có rất nhiều lợi ích, nhưng nấm cũng có không ít tác hại. Ngoài mầm bệnh và chất độc, nấm còn có thể là những kẻ phá hoại ghê gớm. Dưới điều kiện độ ẩm thích hợp, nấm mốc sẽ phát triển và sinh sôi trong các căn nhà. Chúng tiết ra các enzym và acid để phân huỷ các chất hữu cơ, do đó nên chúng có thể phá hoại áo quần, tranh vẽ, phim ảnh, đồ da, sáp, chất cách điện trên dây điện hoặc dây cáp, các chất phủ máy ảnh, máy quay phim và cả chất khởi động máy bay[27]. Chúng là nguyên nhân phổ biến gây thối rữa thức ăn dự trữ, tạo ra những sản phẩm độc hại cho con người và làm suy giảm chất lượng không khí trong nhà. Người ta ước tính có khoảng 40% gia đình ở Mỹ có vấn đề với nấm mốc[109].
Bởi tính năng phân giải cellulose và lignin, nên nhiều loại nấm có thể phá huỷ hay làm mục gỗ ở nhà cửa và công trình xây dựng, gây thiệt hại lớn về kinh tế hàng năm. Có ba dạng nấm làm mục gỗ chính: mục khô (dry rot), mục ẩm (wet rot) và mục mềm (soft rot)[110] dựa theo đặc điểm của gỗ bị mục. Ngoài ra còn có mục nâu (brown rot) - chuyên tấn công và phá hủy các loại quả hạch, như đào, lê, táo, mận[111] - và mục trắng (white rot), dựa theo màu sắc gỗ mục. Những loài nấm phá huỷ gỗ phổ biến nhất là Serpula lacrymans, Coniophora putana, chi Antrodia, Phellinus Contiguous, chi Asterostroma...[110] Để nhằm ngăn chặn quá trình này, một trong số các phương pháp là sử dụng điều khiển sinh học như dùng thông Pinus radiata hay kể cả loại nấm như Phlebiopsis gigantea[112].
Nguồn gốc và phân loại
[sửa | sửa mã nguồn]Trong một thời gian dài, các nhà phân loại học đã xếp nấm là thành viên của giới Thực vật. Sự phân loại này chủ yếu được dựa trên sự tương đồng trong cách sống giữa nấm và thực vật: cả nấm và thực vật chủ yếu đều không di động, hình thái và môi trường sống có nhiều điểm giống nhau (nhiều loài phát triển trên đất, một số loại nấm quả thể giống thực vật như rêu). Thêm nữa, cả hai đều có thành tế bào, điều mà giới Động vật không có. Tuy nhiên, hiện nay nấm lại được công nhận là một giới riêng biệt, khác biệt hẳn với thực vật hay động vật, chúng đã tách ra và xuất hiện xấp xỉ hơn một tỷ năm trước[113]. Nhiều nghiên cứu đã chỉ ra những sự giống và khác về đặc điểm hình thái, sinh hóa và di truyền giữa Nấm và các giới khác. Vì những lý do đó, nấm đã được đặt vào giới riêng của mình.
Lịch sử tiến hóa
[sửa | sửa mã nguồn]
Những cơ thể đầu tiên mang những đặc trưng của nấm xuất hiện vào khoảng 1200 triệu năm (Ma) trước đây, ở Liên đại Nguyên Sinh[115]. Tuy nhiên, những hóa thạch nấm thì không phổ biến cho đến đầu kỷ Devon, khi chúng xuất hiện ở đá Rhynie[116].
Cho dù được xếp cùng với thực vật trước kia, nhưng hiện nay nấm được cho là có quan hệ gần gũi với động vật hơn thực vật, và chúng cùng động vật được xếp vào thành các nhánh của Opisthokonta[117]. Ở Đại Cổ Sinh, nấm xuất hiện trong môi trường nước nước, bao gồm những sinh vật tương tự có mang tiên mao (flagella) tương tự như nấm Chytrid[118]. Những ghi nhận về hóa thạch đầu tiên của nấm rất rời rạc và ít ỏi. Những loài nấm bắt đầu xâm lấn mặt đất trong kỷ Cambri, một thời gian dài trước thực vật[116]. Những lớp nấm hiện đại xuất hiện ở cuối kỷ Than Đá (phân kỷ Pennsylvania)[119]. "Fungal spike", một cum từ được đặt ra bởi các nhà cổ sinh vật học, để nói lên sự phong phú của bào tử nấm trong những trầm tích được tạo ra ngay sau sự kiện tuyệt chủng kỷ Permi-Trias; người ta cho rằng nấm là những dạng sống chủ yếu trong thời kỳ này - khi chúng chiếm đến gần 100% các dữ liệu hóa thạch được tìm thấy [120]. Tuy nhiên thuật ngữ này đã gây ra nhiều tranh cãi: hình dạng của những bào tử nấm (như Reduviasporonites) đôi khi lại tương đồng với bào tử tảo và rất khó để xác định[121], "spike" không xuất hiện trên toàn cầu[122][123] và ở nhiều nơi nó lại không rơi vào giai đoạn Permi-Trias[124].
Những nghiên cứu sử dụng phát sinh loài phân tử củng cố nguồn gốc đơn ngành (monophyletic) của giới Nấm[12]. Hệ thống phân loại nấm thường xuyên thay đổi, đặc biệt bởi những nghiên cứu gần đây dựa trên sự đối chiếu DNA. Những phân tích phát sinh chủng loài hiện hành thường xuyên đánh đổ những hệ thống phân loại mà dựa trên những cách thức cũ và đôi khi kém rõ ràng, ví dụ như dựa trên đặc điểm hình thái và khái niệm loài sinh vật thu được từ những giao phối thực nghiệm[125][126].
Không có bất cứ một hệ thống phân loại nấm ở bậc cao duy nhất nào được công nhận, và những tên gọi thường thay đổi ở mọi cấp độ, kể từ loài trở đi. Dù vậy, những nỗ lực của những nhà nghiên cứu nấm đang được thực hiện để có thể thiết lập và khuyến khích việc sử dụng một danh pháp ổn định và duy nhất[12]. Những loài nấm có thể có nhiều tên khoa học dựa trên vòng đời và phương thức sinh sản. Những trang web như Index Fungorum và ITIS thường xuyên cập nhật những tên gọi mới nhất lẫn tham khảo những tên gọi cũ, nhưng lại không phải lúc nào cũng thống nhất với nhau.
Cây phát sinh
[sửa | sửa mã nguồn]| Unikonta |
| ||||||||||||||||||||||||||||||||||||||||||
Các nhóm phân loại
[sửa | sửa mã nguồn]Các ngành chính của nấm được phân loại chủ yếu dựa trên cấu trúc cơ quan sinh sản hữu tính của chúng. Hiện tại nấm được chia làm 7 ngành:
- Ngành Chytridiomycota hay Chytrid (Nấm roi - nấm trứng): chúng tồn tại rải rác khắp nới trên thế giới. Chytrid sẽ sản sinh ra những bào tử động mà có khả năng di chuyển linh động trong môi trường nước với một tiên mao duy nhất. Vì thế một số nhà phân loại học đã phân loại chúng là những động vật nguyên sinh. Sự phát sinh loài phân tử, dựa trên chuỗi operon rRNA tương ứng ở những tiểu đơn vị ribosome 18S, 28S và 5.8S, cho thấy rằng Chytrid là nhóm nấm căn bản tách ra từ những ngành nấm khác, bao gồm cả bốn ngành chính với những dấu hiệu cho thấy tính cận ngành (paraphyly) hoặc có thể cận ngành của nấm Chytrid[127].
- Ngành Blastocladiomycota trước đây từng được cho là một nhánh phân loại của Chytridiomycota. Những dữ liệu phân tử và đặc điểm siêu cấu trúc gần đây đã đưa Blastocladiomycota vào một nhánh riêng giống như với các ngành Zygomycota, Glomeromycota và Dikarya. Lớp nấm Blastocladiomycetes là những sinh vật hoại sinh hoặc ký sinh của tất cả các nhóm sinh vật nhân chuẩn và chúng giảm phân tạo bào tử, không giống với chytrid, họ hàng gần của chúng, là những loài chủ yếu giảm phân tạo hợp tử[118].
- Ngành Neocallimastigomycota đầu tiên cũng đặt vào ngành Chytridiomycota. Những thành viên của ngành nhỏ này là những sinh vật kỵ khí, sống trong hệ thống tiêu hóa của những động vật ăn cỏ lớn và cũng có thể sống ở môi trường nước và mặt đất. Chúng không có ty thể nhưng lại chứa những hydrogenosome là nguồn gốc của ty thể. Giống như chytrid, neocallimastigomycetes có thể tạo ra những bào tử động mà có một hay nhiều tiên mao ở phía sau[12].
- Ngành Zygomycota (nấm tiếp hợp) có hai lớp: Zygomycetes và Trichomycetes. Chúng sinh sản hữu tính với những bào tử giảm phân được gọi là bào tử tiếp hợp và vô tính với túi bào tử. Loài mốc bánh mỳ đen (Rhizopus stolonifer) là loài phổ biến thuộc ngành này, một chi khác là Pilobolus, có khả năng bắn ra bào tử xa đến vài mét trong không khí. Những chi liên quan đến y học bao gồm Mucor, Rhizomucor và Rhizopus. Những nghiên cứu phát sinh loài phân tử đã chỉ ra rằng Nấm tiếp hợp là nhóm đa ngành và có thể có cận ngành trong nhóm phân loại này[128].
- Những thành viên của ngành Glomeromycota là những nấm tạo ra nấm rễ mút phân nhánh (arbuscular mycorrhizae) ở thực vật bậc cao. Sự cộng sinh này đã có từ cổ đại, với những bằng chứng cho thấy đã có từ 400 triệu năm về trước[24].
- Phân giới Dikarya bao gồm hai ngành Ascomycota và Basidiomycota khi cả hai ngành đều có nhân kép, chúng có thể dạng sợi hoặc đơn bào, nhưng không bao giờ có lông roi. Dikarya được gọi là "Nấm bậc cao", cho dù có nhiều loài sinh sản vô tính được phân loại vào nấm mốc trong các tài liệu trước đây[12]. Deuteromycota (Nấm bất toàn), trước đây từng được coi là một ngành của Nấm, nhưng hiên nay chỉ được sử dụng để chỉ những loại nấm sinh sản vô tính trong Dikarya.
- Ngành Ascomycota (nấm túi hay nấm nang), là nhóm phân loại đông nhất trong Eumycota (Nấm thật). Chúng tạo ra những bào tử giảm phân gọi là bào tử nang, mà được chứa trong một cấu trúc đặc biệt có dạng giống túi gọi là nang (ascus). Ngành này bao gồm nấm nhăn (moscela), vài loại nấm lớn và nấm cục, những nấm men đơn bào (như các chi Saccharomyces, Kluyveromyces, Pichia và Candida) và nhiều nấm sợi sống hoại sinh, ký sinh và cộng sinh. Nhiều loài nấm nang chỉ trải qua quá trình sinh sản vô tính (ở nấm gọi là anamorph), tuy nhiên, những dữ liệu phân tử đã giúp nhận dạng được những giai đoạn hữu tính (teleomorph) gần nhất của chúng ở nấm nang. Bởi những sản phẩm của quá trình giảm phân được chứa trong nang nấm, nên vài loài nấm nang (như Neurospora crassa) được sử dụng để giải thích những nguyên lý của di truyền học.
- Ngành Basidiomycota (Nấm đảm), sản xuất ra những bào tử đảm chứa trong những thân hình dùi gọi là đảm. Đa phần những loài nấm lớn đều thuộc ngành này, cũng như nấm than hay nấm gỉ sắt là những mầm bệnh chính ở ngũ cốc. Những loài nấm đảm quan trọng khác bao gồm nấm Ustilago maydis gây bệnh cho ngô, chi nấm cộng sinh Malassezia gây nên gàu ở người, và nấm cơ hội gây bệnh viêm màng não, Cryptococcus neoformans.
- Một số hình ảnh về các ngành Nấm
-
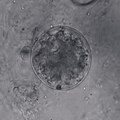 Tế bào nấm thuộc ngành Chytridiomycota
Tế bào nấm thuộc ngành Chytridiomycota -
 Sợi nấm rễ mút phân nhánh nhìn dưới kính hiển vi.
Sợi nấm rễ mút phân nhánh nhìn dưới kính hiển vi. -
 Phycomyces thuộc Zygomycota mọc lên từ một miếng cá
Phycomyces thuộc Zygomycota mọc lên từ một miếng cá -
 Nấm bệnh Mucor mucedo (thuộc Zygomycota) trên thực vật
Nấm bệnh Mucor mucedo (thuộc Zygomycota) trên thực vật -
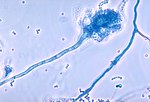 Nấm Aspergillus fumigatus với các bào tử nang
Nấm Aspergillus fumigatus với các bào tử nang -
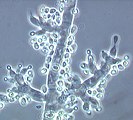 Trichoderma fertile thuộc Ascomycota
Trichoderma fertile thuộc Ascomycota -
 Pneumocystis carinii gây bệnh viêm phổi ở người
Pneumocystis carinii gây bệnh viêm phổi ở người -
 Cryptococcus neoformans với lớp màng polysaccarit ở nhiệt độ 37 °C
Cryptococcus neoformans với lớp màng polysaccarit ở nhiệt độ 37 °C -
 Nấm mồng gà (Cantharellus) thuộc Basidiomycota
Nấm mồng gà (Cantharellus) thuộc Basidiomycota -
 Nấm linh chi (Ganoderma lucidum) được sử dụng trong y học phương Đông từ lâu đời
Nấm linh chi (Ganoderma lucidum) được sử dụng trong y học phương Đông từ lâu đời -
 Humidicutis lewelliniae thuộc Basidiomycota
Humidicutis lewelliniae thuộc Basidiomycota











Mối quan hệ với những sinh vật giống nấm khác
[sửa | sửa mã nguồn]Bởi vì có những sự tương đồng giữa hình thái và vòng đời, nên nấm nhầy (myxomycetes) và mốc nước (oomycetes) trước đây đã được đặt vào giới Nấm. Tuy nhiên, không giống nấm thật (Eumycota), thành tế bào của những sinh vật này có chứa cellulose và thiếu kitin. Nấm nhầy cũng là Unikont giống như nấm, nhưng lại thuộc giới Amoebozoa. Mốc nước là những Bikont lưỡng bội thuộc giới Chromalveolate. Cả mốc nước và nấm nhầy đều không có quan hệ gần gũi với nấm thật và vì thế những nhà phân loại học hiện nay đã không xếp chúng vào giới Nấm. Dù vậy, ngành học nghiên cứu về myxomycete và oomycete vẫn thường được xếp nào nấm học trong những sách giáo khoa và tài liệu nghiên cứu sơ cấp.
Trước đây đã có giả thuyết cho rằng Nucleariid có thể là nhánh chị em với mốc nước, và vì thế cho rằng có thể thuộc giới Nấm[129]. Tuy nhiên, hiện tại thì Nucleariid vẫn đang nằm trong giới Choanozoa.
Xem thêm
[sửa | sửa mã nguồn]Chú thích
[sửa | sửa mã nguồn]- ^ Moore RT. (1980). “Taxonomic proposals for the classification of marine yeasts and other yeast-like fungi including the smuts”. Botanica Marine. 23: 361–373.
- ^ The classification system presented here is based on the 2007 phylogenetic study by Hibbett.
- ^ Vaupotic T, Veranic P, Jenoe P, Plemenitas A. (2008). “Mitochondrial mediation of environmental osmolytes discrimination during osmoadaptation in the extremely halotolerant black yeast Hortaea werneckii”. Fungal Genetics and Biology. 45 (6): 994–1007. doi:10.1016/j.fgb.2008.01.006. ISSN 1087-1845. PMID 18343697.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Dadachova E, Bryan RA, Huang X, Moadel T, Schweitzer AD, Aisen P, Nosanchuk JD, Casadevall A. (2007). “Ionizing radiation changes the electronic properties of melanin and enhances the growth of melanized fungi”. PLoS ONE. 2 (5): e457. Bibcode:2007PLoSO...2..457D. doi:10.1371/journal.pone.0000457. PMC 1866175. PMID 17520016.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Raghukumar C, Raghukumar S. (1998). “Barotolerance of fungi isolated from deep-sea sediments of the Indian Ocean”. Aquatic Microbial Ecology. 15 (2): 153–163. doi:10.3354/ame015153.
- ^ Le Calvez T, Burgaud G, Mahé S, Barbier G, Vandenkoornhuyse P. (2009). “Fungal diversity in deep sea hydrothermal ecosystems”. Applied and Environmental Microbiology. 75 (20): 6415–6421. doi:10.1128/AEM.00653-09. PMC 2765129. PMID 19633124.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Hawksworth DL (2006). “The fungal dimension of biodiversity: magnitude, significance, and conservation”. Mycol. Res. 95: 641–655.
- ^ This estimation is determined by combining the species count for each phyla, based on values obtained from the 10th edition of the Dictionary of the Fungi (Kirk., 2008): Ascomycota, 64163 species (p. 55); Basidiomycota, 31515 (p. 78); Blastocladiomycota, 179 (p. 94); Chytridiomycota, 706 (p. 142); Glomeromycota, 169 (p. 287); Microsporidia, >1300 (p. 427); Neocallimastigomycota, 20 (p. 463).
- ^ Mueller G.M., Schmit J.P. (2006). “Fungal biodiversity: what do we know? What can we predict?”. Biodivers Conserv. 16: 1–5. doi:10.1007/s10531-006-9117-7.
- ^ Alexopoulos C.J., Mims C.W., Blackwell M. (1996). Introductory Mycology. John Wiley and Sons. ISBN 0471522295.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Meredith Blackwell (ngày 14 tháng 2 năm 2005). “Eumycota: mushrooms, sac fungi, yeast, molds, rusts, smuts, etc.” (bằng tiếng Anh). Bản gốc lưu trữ ngày 25 tháng 1 năm 2021. Truy cập ngày 9 tháng 8 năm 2008. Đã định rõ hơn một tham số trong author-name-list parameters (trợ giúp)
- ^ a b c d e Hibbett D.S., và ctv. (2007). “A higher level phylogenetic classification of the Fungi”. Mycol. Res. 111 (5): 509–547. doi:10.1016/j.mycres.2007.03.004.
- ^ Lindahl B.D., Ihrmark K., Boberg J., Trumbore S.E., Högberg P., Stenlid J., Finlay R.D. (2007). “Spatial separation of litter decomposition and mycorrhizal nitrogen uptake in a boreal forest”. New Phytol. 173: 611–620. doi:10.1111/j.1469-8137.2006.01936.x. PMID 17244056.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Barea J.M., Pozo M.J., Azcón R., Azcón-Aguilar C. (2005). “Microbial co-operation in the rhizosphere”. J. Exp. Bot. 56: 1761–1778. doi:10.1093/jxb/eri197. PMID 15911555.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Aanen D.K. (2006). “As you reap, so shall you sow: coupling of harvesting and inoculating stabilizes the mutualism between termites and fungi”. Biol Lett. 2: 209–212. doi:10.1098/rsbl.2005.0424. PMID 17148364.
- ^ Nikoh N., Fukatsu T. (2000). “Interkingdom host jumping underground: phylogenetic analysis of entomoparasitic fungi of the genus Cordyceps”. Mol Biol Evol. 17: 2629–2638. PMID 10742053.
- ^ Perotto S., Bonfante P. (1997). “Bacterial associations with mycorrhizal fungi: close and distant friends in the rhizosphere”. Trends Microbiol. 5: 496–501. doi:10.1016/S0966-842X(97)01154-2. PMID 9447662.
- ^ Arnold A.E., Mejía L.C., Kyllo D., Rojas E.I., Maynard Z., Robbins N., Herre E.A. (2003). “Fungal endophytes limit pathogen damage in a tropical tree”. Proc. Natl. Acad. Sci. USA. 100: 15649–15654. doi:10.1073/pnas.2533483100. PMID 14671327.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ a b Paszkowski U. (2006). “Mutualism and parasitism: the yin and yang of plant symbioses”. Curr Opin Plant Biol. 9: 364–370. doi:10.1016/j.pbi.2006.05.008. PMID 16713732.
- ^ a b Hube B. (2004). “From commensal to pathogen: stage- and tissue-specific gene expression of Candida albicans”. Curr Opin Microbiol. 7: 336–341. doi:10.1016/j.mib.2004.06.003. PMID 15288621.
- ^ Volk, Tom. “Tom Volk's Fungi FAQ”. Truy cập ngày 12 tháng 8 năm 2008.
- ^ Wong, George. “Symbiosis: Mycorrhizae and Lichens”. Truy cập ngày 12 tháng 8 năm 2008.
- ^ Knowledge of nitrogen transfer between plants and beneficial fungi expands, southwestfarmpress.com, 10-6-2005, truy cập 9-8-2008.
- ^ a b Remy W., Taylor T.N., Hass H., Kerp H. (1994). “4-hundred million year old vesicular-arbuscular mycorrhizae”. Proc. Natl. Acad. Sci. 91: 11841–11843. doi:10.1073/pnas.91.25.11841. PMID 11607500.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ van der Heijden M.G., Streitwolf-Engel R., Riedl R., Siegrist S., Neudecker A., Ineichen K., Boller T., Wiemken A., Sanders I.R. (2006). “The mycorrhizal contribution to plant productivity, plant nutrition and soil structure in experimental grassland”. New Phytol. 172: 739–752. doi:10.1111/j.1469-8137.2006.01862.x. PMID 17096799.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Selosse M.A., Richard F., He X., Simard S.W. (2006). “Mycorrhizal networks: des liaisons dangereuses?”. Trends Ecol Evol. 21: 621–628. doi:10.1016/j.tree.2006.07.003. PMID 16843567.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ a b c d W.D.Phillips & T.J. Chilton (1991). “52”. Sinh học (A. Level Biology). 2. Việt Nam: Nhà xuất bản Giáo dục. tr. 217–228. ISBN 9780199140893.
- ^ Irwin M. Brodo & Sylvia Duran Sharnoff (2001). Lichens of North America. Nhà in Đại học Yale. ISBN 0300082495.Quản lý CS1: sử dụng tham số tác giả (liên kết)
- ^ Jocelyn G. Millar & Kenneth F. Haynes (1998). Methods in Chemical Ecology. Springer. tr. 163. ISBN 0412080419.Quản lý CS1: sử dụng tham số tác giả (liên kết)
- ^ Fungi and Insect Symbiosis www.botany.hawaii.edu. Truy cập 10-8-2008.
- ^ Pascal Jouquet, Virginie Tavernier, Luc Abbadie & Michel Lepage. Nests of subterranean fungus-growing termites (Isoptera, Macrotermitinae) as nutrient patches for grasses in savannah ecosystems. African Journal of Ecology. 2005. Vol 43, 191–196
- ^ Talbot N.J. (2003). “On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea”. Annu Rev Microbiol. 57: 177–202. doi:10.1146/annurev.micro.57.030502.090957. PMID 14527276.
- ^ Paoletti M., Buck K.W., Brasier C.M. (2006). “Selective acquisition of novel mating type and vegetative incompatibility genes via interspecies gene transfer in the globally invading eukaryote Ophiostoma novo-ulmi”. Mol Ecol. 15: 249–262. doi:10.1111/j.1365-294X.2005.02728.x. PMID 16367844.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Gryzenhout M., Wingfield B.D., Wingfield M.J. (2006). “New taxonomic concepts for the important forest pathogen Cryphonectria parasitica and related fungi”. FEMS Microbiol Lett. 258: 161–172. doi:10.1111/j.1574-6968.2006.00170.x. PMID 16640568.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Nielsen K., Heitman J. (2007). “Sex and virulence of human pathogenic fungi”. Adv Genet. 57: 143–173. doi:10.1016/S0065-2660(06)57004-X. PMID 17352904.
- ^ Brakhage A.A. (2005). “Systemic fungal infections caused by Aspergillus species: epidemiology, infection process and virulence determinants”. Curr. Drug Targets. 6: 875–886. doi:10.2174/138945005774912717. PMID 16375671.
- ^ Kauffman C.A. (2007). “Histoplasmosis: a clinical and laboratory update”. Clin Microbiol Rev. 20: 115–132. doi:10.1128/CMR.00027-06. PMID 17223625.
- ^ Cushion M.T., Smulian A.G., Slaven B.E., Sesterhenn T., Arnold J., Staben C., Porollo A., Adamczak R., Meller J. (2007). “Transcriptome of Pneumocystis carinii during Fulminate Infection: Carbohydrate Metabolism and the Concept of a Compatible Parasite”. PLoS ONE. 2: e423. doi:10.1371/journal.pone.0000423. PMID 17487271.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Howard E. Gendelman, Igor Grant, Ian P. Everall, Stuart A. Lipton, Susan Swindells (2005). The Neurology of AIDS. Oxford University Press. tr. 503. ISBN 0198526105.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Kevin B. Frey, Paul Price, John Paul Price (2003). microbiology for surgical technologists. Thomson Delmar Learning. tr. 366–367. ISBN 0766826996.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ a b Kevin Kavanagh (2005). Fungi: Biology and Applications. John Wiley and Sons. tr. 173-187. ISBN 0470867019.
- ^ Michael Breitenbach, Reto Crameri, Samuel B. Lehrer (2002). Fungal Allergy and Pathogenicity. Karger Publishers. ISBN 380557391X.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ a b John H. Krouse, Stephen J. Chadwick, Bruce R. Gordon, M. Jennifer Derebery (2001). Allergy and Immunology: An Otolaryngic Approach. Lippincott Williams & Wilkins. tr. 43–46. ISBN 078172628X.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ a b Richard F. Lockey, Samuel C. Bukantz, Jean Bousquet (2004). Allergens and Allergen Immunotherapy. Informa Health Care. tr. 223-259. ISBN 0824756509.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ William Briner, American College of Sports Medicine (2006). Action Plan for Allergies. Human Kinetics. tr. 150–154. ISBN 0736062793.
- ^ a b c Nguyễn Lân Dũng (ngày 11 tháng 6 năm 2006). “Vi nấm (Microfungi)”. Chương trình Vi sinh vật. Vietsciences. Truy cập ngày 10 tháng 8 năm 2008. Đã định rõ hơn một tham số trong author-name-list parameters (trợ giúp)
- ^ Ying Yang (ngày 27 tháng 3 năm 2007). “Evolution of nematode-trapping cells of predatory fungi of the Orbiliaceae based on evidence from rRNA-encoding DNA and multiprotein sequences” (pdf) (bằng tiếng Anh). Truy cập ngày 10 tháng 8 năm 2008. Đã định rõ hơn một tham số trong author-name-list parameters (trợ giúp)
- ^ Sigee, D. C. (2005). Freshwater Microbiology. John Wiley and Sons. tr. 389. ISBN 0471485292, 9780471485292 Kiểm tra giá trị
|isbn=: ký tự không hợp lệ (trợ giúp). - ^ Leslie, Anne R. (1994). Handbook of Integrated Pest Management for Turf and Ornamentals. CRC Press. tr. 495–497. ISBN 0873713508, 9780873713504 Kiểm tra giá trị
|isbn=: ký tự không hợp lệ (trợ giúp). - ^ Decker, Heinz (1989). Plant nematodes and their control (phytonematology). BRILL. tr. 114–115. ISBN 9004089225, 9789004089228 Kiểm tra giá trị
|isbn=: ký tự không hợp lệ (trợ giúp). - ^ Maurice Langeron & R. Vanbreuseghem (1965). Outline of Mycology. C.C. Thomas. tr. 109.Quản lý CS1: sử dụng tham số tác giả (liên kết)
- ^ Pereira J.L., Noronha E.F., Miller R.N., Franco O.L. (2007). “Novel insights in the use of hydrolytic enzymes secreted by fungi with biotechnological potential”. Lett Appl Microbiol. 44: 573–581. doi:10.1111/j.1472-765X.2007.02151.x. PMID 17576216.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Schaller M., Borelli C., Korting H.C., Hube B. (2007). “Hydrolytic enzymes as virulence factors of Candida albicans”. Mycoses. 48: 365–377. doi:10.1111/j.1439-0507.2005.01165.x. PMID 16262871.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Farrar J.F. (1985). “Carbohydrate metabolism in biotrophic plant pathogens”. Microbiol Sci. 2: 314–317. PMID 3939987.
- ^ Marzluf G.A. (1981). “Regulation of nitrogen metabolism and gene expression in fungi”. Microbiol Rev. 45: 437–461. PMID 6117784.
- ^ Heynes M.J. (1994). “Regulatory circuits of the amdS gene of Aspergillus nidulans”. Antonie Van Leeuwenhoek. 65: 179–782. doi:10.1007/BF00871944. PMID 7847883.
- ^ a b Dadachova E., Bryan R.A., Huang X., Moadel T., Schweitzer A.D., Aisen P., Nosanchuk J.D., Casadevall A. (2007). “Ionizing radiation changes the electronic properties of
melanin and enhances the growth of melanized fungi”. PLoS ONE. 2: e457. doi:10.1371/journal.pone.0000457. PMID 17520016. line feed character trong
|title=tại ký tự số 56 (trợ giúp)Quản lý CS1: nhiều tên: danh sách tác giả (liên kết) - ^ Kulp, Karel (2000). Handbook of Cereal Science and Technology. CRC Press. ISBN 0824782941.
- ^ Piskur J., Rozpedowska E., Polakova S., Merico A., Compagno C. (2006). “How did Saccharomyces evolve to become a good brewer?”. Trends Genet. 22: 183–186. doi:10.1016/j.tig.2006.02.002. PMID 16499989.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Ai Leng Teoh & Gillian Heard and Julian Cox (ngày 1 tháng 9 năm 2004). “Yeast ecology of Kombucha fermentation”. International Journal of Food Microbiology. 95 (2): 119–126. doi:10.1016/j.ijfoodmicro.2003.12.020. Truy cập ngày 24 tháng 12 năm 2006.Quản lý CS1: sử dụng tham số tác giả (liên kết)
- ^ Hickling, S., Hung, J., Knuiman, M., Jamrozik, K., McQuillan, B., Beilby, J., Thompson, P. (2005). “Impact of voluntary folate fortification on plasma homocysteine and serum folate in Australia from 1995 to 2001: a population based cohort study”. Journal of Epidemiology & Community Health. 59: 371–376.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ a b c Nguyễn Lân Dũng (ngày 26 tháng 9 năm 2005). “Jokichi Takamine (1854-1922) Người cha Nhật Bản của Công nghệ sinh học Hoa Kỳ, cha đẻ Adrénaline, khám phá Aspergillus oryzae”. Vietsciences. Truy cập ngày 11 tháng 8 năm 2008.
- ^ Kitamoto N., Yoshino S., Ohmiya K., Tsukagoshi N. (1999). “Sequence analysis, overexpression, and antisense inhibition of a beta-xylosidase gene, xylA, from Aspergillus oryzae KBN616”. Appl. Env. Microbiol. 65: 20–24. PMID 9872754.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Dilip K. Arora & George G. Khachatourians, ScienceDirect (2003). Applied Mycology and Biotechnology. Elsevier. tr. 134. ISBN 0444510303.Quản lý CS1: sử dụng tham số tác giả (liên kết)
- ^ a b c d Dilip K. Arora & P. D. Bridge, Deepak Bhatnagar (2004). Fungal Biotechnology in Agricultural, Food, and Environmental. CRC Press. tr. 228–235. ISBN 0824747704.Quản lý CS1: sử dụng tham số tác giả (liên kết)
- ^ Shawn M. Talbott & Kerry Hughes (2007). The Health Professional's Guide to Dietary Supplements. Lippincott Williams & Wilkins. tr. 227. ISBN 0781746728.Quản lý CS1: sử dụng tham số tác giả (liên kết)
- ^ Nina Gunde-Cimerman & Aharon Oren, Ana Plemenitaš (2005). Adaptation to Life at High Salt Concentrations in Archaea, Bacteria, and Eukarya. Springer. tr. 445. ISBN 1402036329.Quản lý CS1: sử dụng tham số tác giả (liên kết)
- ^ Vai trò hữu dụng của nấm trong chế biến thực phẩm, Nguyễn Văn Thành, cập nhật 22/7/2009
- ^ Didi Emmons & Melissa Sweet (1997). Vegetarian Planet: 350 Big-flavor Recipes for Out-of-this-world Food Every Day. Harvard Common Press. tr. 141-142. ISBN 1558321152.Quản lý CS1: sử dụng tham số tác giả (liên kết)
- ^ “Bringing Mushrooms Out of the Dark”. MSNBC. ngày 18 tháng 4 năm 2006. Truy cập ngày 6 tháng 8 năm 2007.
- ^ Genders, Roy (1969). Mushroom Growing for Everyone. London: Faber. tr. 121. ISBN 0-571-08992-5.
- ^ (tiếng Ý)Cappelli, Alberto (1984). Fungi Europaei:Agaricus. Saronno, Italy: Giovanna Biella. tr. trang 123-25.
- ^ Roody, William C. (2003). Mushrooms of West Virginia and the Central Appalachians. Nhà in Đại học Kentucky. tr. 4. ISBN 0813190398.
- ^ On the Trail of the Death Cap Mushroom Richard Harris, NPR, 8-2-2007. Truy cập 11-8-2008.
- ^ Leathem A.M., Dorran T.J. (2007). “Poisoning due to raw Gyromitra esculenta (false morels) west of the Rockies”. CJEM. 9: 127–130. PMID 17391587.
- ^ Karlson-Stiber C., Persson H. (2003). “Cytotoxic fungi--an overview”. Toxicon. 42: 339–349. doi:10.1016/S0041-0101(03)00238-1. PMID 14505933.
- ^ Mythology and Folklore of Fly Agaric Lưu trữ 2008-07-23 tại Wayback Machine Paul Kendall, Trees for Life. Truy cập 11-8-2008.
- ^ Phân biệt nấm ăn và nấm độc, Nguyễn Văn Chất (theo báo HàNộimới, Thanh Niên, 6-11-2004. Truy cập 11-8-2008
- ^ Nấm ăn và nấm độc Lưu trữ 2016-03-07 tại Wayback Machine, tapchivhntanuong.com.vn, Hương Liên, 15-6-2007. Truy cập 11-8-2008
- ^ Nguyễn Lân Dũng, Dương Văn Hợp, Phạm Thế Hải (ngày 3 tháng 10 năm 2005). “Thần dược Đông Trùng Hạ Thảo”. Vietsciences. Truy cập ngày 12 tháng 8 năm 2008. Chú thích có tham số trống không rõ:
|accessmonthday=(trợ giúp)Quản lý CS1: nhiều tên: danh sách tác giả (liên kết) - ^ Chưa có cơ sở để nói nấm cổ linh chi chữa được bệnh, VnExpress, 21-5-2003. Truy cập 12-8-2008
- ^ Francisco A. Moreno, Pedro Delgado, Alan J. Gelenberg. “Effects of Psilocybin in Obsessive-Compulsive Disorder” (bằng tiếng Anh). MAPS. Bản gốc lưu trữ ngày 13 tháng 2 năm 2012. Truy cập ngày 12 tháng 8 năm 2008. Chú thích có tham số trống không rõ:
|accessmonthday=(trợ giúp)Quản lý CS1: nhiều tên: danh sách tác giả (liên kết) - ^ Hallucinogens and Obsessive-Compulsive Disorder Lưu trữ 2010-03-29 tại Wayback Machine, Daniel M. Perine. Truy cập 12-08-2008
- ^ LSD & Psilocybin Research for Cluster Headache Treatment Lưu trữ 2009-01-27 tại Wayback Machine, MAPS. Truy cập 12-8-2008
- ^ Wilson, John Rowan (1964). The Mind. Time, inc. tr. 157.
- ^ Stephen Wrigley, Royal Society of Chemistry (Great Britain), M. Hayes, R. Thomas, E. J. T. Chrystal (2000). Biodiversity: New Leads for the Pharmaceutical and Agrochemical Industries. Royal Society of Chemistry. tr. 4. ISBN 0854048308.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Takuji Hara (2003). Innovation in the Pharmaceutical Industry: The Process of Drug Discovery and Development. Edward Elgar Publishing. tr. 145. ISBN 1843760509.
- ^ a b Oliver Zerbe, Raimund Mannhold, Hugo Kubinyi, Gerd Folkers (2006). BioNMR in Drug Research. Wiley-VCH. tr. 22–24. ISBN 3527605428.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Zhiqiang An (2004). Handbook of Industrial Mycology. CRC Press. tr. 667–688. ISBN 082475655X.
- ^ Margaret A. Powers (1996). Handbook of Diabetes Medical Nutrition Therapy. Jones & Bartlett Publishers. tr. 81. ISBN 0834206315.
- ^ a b c d Zhiqiang An, trang 1-26
- ^ “Trichoderma spp., including T. harzianum, T. viride, T. koningii, T. hamatum and other spp. Deuteromycetes, Moniliales (asexual classification system)”. Biological Control: A Guide to Natural Enemies in North America. Truy cập ngày 18 tháng 8 năm 2008.
- ^ A. Altman, Rita R. Colwell (1997). Agricultural Biotechnology. CRC Press. tr. 357. ISBN 0824794397.
- ^ Stamets, Paul (2005). Mycelium Running: How Mushrooms Can Help Save the World. Ten Speed Press. tr. 51. ISBN 1580085792.
- ^ Stamets, Paul, 51
- ^ Douglas, Michael S., 17/4/1995. [1]Lưu trữ 2012-01-19 tại Wayback Machine Bioremediation of Contaminated Soils Using the White Rot Fungus Phanerochaete Chrysosporium. DTIC.
- ^ Stamets, Paul, 109
- ^ BBC. (2008). Fungi to fight 'toxic war zones'. Truy cập 12-08-2008
- ^ López-Gómez J, Molina-Meyer M (2006). “The competitive exclusion principle versus biodiversity through competitive segregation and further adaptation to spatial heterogeneities”. Theor Popul Biol. 69: 94–109. doi:10.1016/j.tpb.2005.08.004. PMID 16223517.
- ^ Stamets, Paul, 113
- ^ Setting the Stage To Screen Biocontrol Fungi Hank Becker, Tháng 7 năm 1998. Truy cập 13-08-2008.
- ^ [2]Lưu trữ 2011-12-29 tại Wayback Machine Whey-based fungal microfactory technology for enhanced biological pest management using fungi Todd. S. Keiller, Technology Transfer, University of Vermont. Truy cập 12-08-2008.
- ^ Deshpande MV. (1999). “Mycopesticide production by fermentation: potential and challenges”. Crit Rev Microbiol. 25: 229–243. doi:10.1080/10408419991299220. PMID 10524330.
- ^ Thomas MB, Read AF. (2007). “Can fungal biopesticides control malaria?”. Nat Rev Microbiol. 5: 377–383. doi:10.1038/nrmicro1638. PMID 17426726.
- ^ I.M. Hall & P.H. Dunn, Entomophthorous Fungi Parasitic on the Spotted Alfalfa Aphid, Hilgardia, Tháng 12 năm 1957.
- ^ Harman, G.E. (2006). “Overview of mechanisms and uses of Trichoderma spp”. Phytopathology. 96: 190–194. doi:10.1094/PHYTO-96-0190.
- ^ Bush LP, Wilkinson HH, Schardl CL. (1997). “Bioprotective Alkaloids of Grass-Fungal Endophyte Symbioses”. Plant Physiol. 114: 1–7. PMID 12223685.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Bouton JH, Latch GCM, Hill NS, Hoveland CS, McCannc MA, Watson RH, Parish JA, Hawkins LL, Thompson FN (2002). “Use of nonergot alkaloid-producing endophytes for alleviating tall fescue toxicosis in sheep”. Agron. J. 94: 567–574. http://web.archive.org/web/20030708205426/http://agron.scijournals.org/cgi/content/full/94/3/567.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Niemeier, R. Todd, Sivasubramani, Satheesh K., Reponen, Tiina and Grinshpun, Sergey A., (2006) "Assessment of Fungal Contamination in Moldy Homes: Comparison of Different Methods", Journal of Occupational and Environmental Hygiene, 3:5, 262-273 [1]
- ^ a b R. O. Heckroodt (2002). Guide to the Deterioration and Failure of Building Materials. Thomas Telford. tr. 88–94. ISBN 0727731726.
- ^ Brown Rot of Stone Fruits, Michael A. Ellis, Ohio State University Extension Fact Sheet. Truy cập 13-08-2008.
- ^ Biological control of sapwood staining fungi Lưu trữ 2008-08-03 tại Wayback Machine, University of Minnesota. Truy cập 13-08-2008
- ^ Bruns T. (2006). “Evolutionary biology: a kingdom revised”. Nature. 443: 758–761. PMID 17051197.
- ^ The Humongous Fungus--Ten Years Later ở trang Đại học Wisconsin. Truy cập 17-8-2008
- ^ Butterfield, N.J. (2005). “Probable Proterozoic fungi”. Paleobiology. 31 (1): 165–182.
- ^ a b Brundrett, M.C. (2002). “Coevolution of roots and mycorrhizas of land plants”. New Phytologist. 154 (2): 275–304. doi:10.1046/j.1469-8137.2002.00397.x.
- ^ P. Sitte, H. Ziegler, F. Ehrendorfer (1991). Strasburger Lehrbuch der Botanik (Textbook of Botany) (ấn bản thứ 33). Urban & Fischer. ISBN 3437204475.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ a b James TYyear= 2006. “Reconstructing the early evolution of Fungi using a six-gene phylogeny”. Nature. 443: 818–822. doi:10.1038/nature05110. PMID 17051209.
- ^ Meredith Blackwell, Rytas Vilgalys, Timothy Y. James, and John W. Taylor. “Fungi Eumycota: mushrooms, sac fungi, yeast, molds, rusts, smuts, etc”. The Tree of Life Web Project. Bản gốc lưu trữ ngày 24 tháng 9 năm 2008. Truy cập ngày 15 tháng 8 năm 2008.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Eshet, Y. (1995) Fungal event and palynological record of ecological crisis and recovery across the Permian-Triassic boundary. Geology, 23, 967-970.
- ^ Foster, C.B. (2002). Stephenson, M.H.; Marshall, C.; Logan, G.A.; Greenwood, P.F. “A Revision Of Reduviasporonites Wilson 1962: Description, Illustration, Comparison And Biological Affinities”. Palynology. 26 (1): 35–58. doi:10.2113/0260035.
- ^ López-Gómez, J. and Taylor, E.L. (2005). “Permian-Triassic Transition in Spain: A multidisciplinary approach”. Palaeogeography, Palaeoclimatology, Palaeoecology. 229 (1–2): 1–2. doi:10.1016/j.palaeo.2005.06.028. Bản gốc lưu trữ ngày 6 tháng 3 năm 2010. Truy cập ngày 15 tháng 8 năm 2008.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Looy, C.V. (2005). Twitchett, R.J.; Dilcher, D.L.; Van Konijnenburg-van Cittert, J.H.A.; Visscher, H. “Life in the end-Permian dead zone”. Proceedings of the National Academy of Sciences. 162 (4): 653–659. doi:10.1073/pnas.131218098. PMID 11427710.
See image 2
- ^ Ward PD, Botha J, Buick R, De Kock MO, Erwin DH, Garrison GH, Kirschvink JL & Smith R (2005). “Abrupt and Gradual Extinction Among Late Permian Land Vertebrates in the Karoo Basin, South Africa”. Science. 307 (5710): 709–714. doi:10.1126/science.1107068.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Palaeos: Fungi. Truy cập 15-08-2008
- ^ David S. Hibbett (ngày 8 tháng 12 năm 2007). “A Higher-Level Phylogenetic Classification of the Fungi by” (PDF). Mycological Research. Clark University. Bản gốc (pdf) lưu trữ ngày 26 tháng 3 năm 2009. Truy cập ngày 15 tháng 8 năm 2008.
- ^ James TY, Letcher PM, Longcore JE, Mozley-Standridge SE, Porter D, Powell MJ, Griffith GW, Vilgalys R. (2006). “A molecular phylogeny of the flagellated fungi (Chytridiomycota) and description of a new phylum (Blastocladiomycota)”. Mycologia. 98: 860–871. doi:10.3852/mycologia.98.6.860. PMID 17486963.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ White MM, James TY, O'Donnell K, Cafaro MJ, Tanabe Y, Sugiyama J. (2006). “Phylogeny of the Zygomycota based on nuclear ribosomal sequence data”. Mycologia. 98: 872–884. doi:10.3852/mycologia.98.6.872. PMID 17486964.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Karl Esser & Paul A. Lemke (1994). The Mycota: A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research. Springer. ISBN 3540580085.Quản lý CS1: sử dụng tham số tác giả (liên kết)
Liên kết ngoài
[sửa | sửa mã nguồn]- Comparative Analysis of Fungal Genomes Lưu trữ 2008-10-05 tại Wayback Machine (at DOE's IMG system)
- Tree of Life web project: Fungi Lưu trữ 2021-01-25 tại Wayback Machine
- Nấm tại Encyclopedia of Life
- Mushroom Observer (mushroomobserver.org), a collaborative fungus recording and identification project
.jpg)
.jpg)









 GIẢM
24%
GIẢM
24%
 GIẢM
22%
GIẢM
22%
 GIẢM
20%
GIẢM
20%


 GIẢM
43%
GIẢM
43%



