Khủng long
| Khủng long | |||
|---|---|---|---|
| Khoảng thời gian tồn tại: (Có lẽ bao quát cả Trias giữa) | |||

| |||
| Một số loài khủng long tiêu biểu. Theo chiều kim đồng hồ từ góc trái trên cùng: Microraptor gui - khủng long chân thú Apatosaurus louisae - khủng long chân thằn lằn Edmontosaurus regalis - khủng long chân chim Triceratops horridus - khủng long mặt sừng Stegosaurus stenops - khủng long phiến sừng Pinacosaurus grangeri - khủng long bọc giáp | |||
| Phân loại khoa học | |||
| Vực: | Eukaryota | ||
| Giới: | Animalia | ||
| Ngành: | Chordata | ||
| nhánh: | Sauropsida | ||
| nhánh: | Archosauria | ||
| nhánh: | Avemetatarsalia | ||
| nhánh: | Ornithodira | ||
| nhánh: | Dinosauromorpha | ||
| nhánh: | Dinosauriformes | ||
| nhánh: | Dracohors | ||
| nhánh: | Dinosauria Owen, 1842 | ||
| Các nhóm lớn | |||
| |||

Khủng long là một nhóm bò sát[phụ chú 1] thuộc nhánh Dinosauria, xuất hiện lần đầu vào kỷ Tam Điệp khoảng 243 - 233,23 triệu năm trước đây, mặc dù nguồn gốc chính xác và các mốc thời gian trong quá trình tiến hóa của chúng hiện vẫn đang tiếp tục được đào sâu nghiên cứu. Kể từ sau sự kiện tuyệt chủng Tam Điệp – Jura (201,3 triệu năm trước), khủng long đã trở thành nhóm động vật có xương sống chiếm ưu thế nhất xuyên suốt qua kỷ Jura cho đến cuối kỷ Phấn Trắng, khi sự kiện tuyệt chủng Phấn Trắng-Cổ Cận (66 triệu năm trước) làm tuyệt chủng hầu hết các nhóm khủng long và 3/4 các loài động vật, thực vật trên Trái Đất (trong đó đa số các loài bò sát khổng lồ bao gồm khủng long bị thiệt hại nặng nề nhất, các loài cá và lưỡng cư ít bị thiệt hại), sự kiện tuyệt chủng này đánh dấu sự kết thúc Đại Trung Sinh và bắt đầu Đại Tân Sinh. Các ghi nhận hóa thạch cho thấy chim đã tiến hóa từ khủng long có lông vũ, tiến hóa từ một nhóm lớn hơn là khủng long chân thú vào thế Jura muộn, và vài loài chim đã sống sót sau sự kiện tuyệt chủng 66 triệu năm trước rồi tiếp tục phát triển đa dạng như hiện nay. Do đó, chim là hậu duệ duy nhất còn sót lại ngày nay của khủng long, và khủng long vẫn chưa tuyệt chủng hoàn toàn như vẫn thường được nói mà một nhánh con cháu của chúng vẫn còn tồn tại đến tận ngày nay. Khủng long vì thế có thể được chia thành hai loại là khủng long phi điểu (tức là tất cả các loại khủng long khác chim) và chim.
Khủng long là một nhóm đa dạng từ phân loại sinh học, hình thái đến sinh thái. Chim, với hơn 10,000 loài còn sinh tồn, là nhóm động vật có xương sống đa dạng nhất ngoài bộ Cá vược. Theo các bằng chứng hóa thạch, các nhà cổ sinh đã nhận ra 500 chi và hơn 1000 loài khủng long phi điểu. Khủng long có mặt ở khắp các châu lục, qua những loài hiện còn (chim) cũng như những hóa thạch còn sót lại. Suốt nửa đầu của thế kỷ XX, trước khi chim được xem là khủng long, hầu hết cộng đồng khoa học tin rằng khủng long là động vật chậm chạp và biến nhiệt. Tuy nhiên hầu hết các nghiên cứu những năm 1970 đã chỉ ra rằng khủng long là động vật hoạt động tích cực với khả năng trao đổi chất cao và thích nghi tốt cho quan hệ xã hội. Một số ăn thực vật, số khác ăn thịt. Có bằng chứng cho thấy tất cả khủng long đều đẻ trứng, và xây tổ là một đặc điểm phụ của tất cả khủng long, cả chim lẫn phi điểu.
Tổ tiên khủng long đi bằng hai chân. Tuy thế rất nhiều chi khủng long đi bằng bốn chân, và một số chi có thể thay đổi giữa 2 dạng. Cấu trúc sừng và mào là phổ biến ở tất cả các nhóm khủng long, và vài nhóm thậm chí còn phát triển các biến đổi bộ xương như giáp mô hoặc gai. Trong khi khủng long hiện đại (chim) khá nhỏ để thuận tiện cho việc bay lượn (tất nhiên vẫn có một vài loài chim không thể bay), nhiều loài khủng long có cơ thể rất lớn. Chi khủng long chân thằn lằn lớn nhất có thể đạt tới 39,7 mét (130 foot) chiều dài và 18 m (59 ft) chiều cao. Dù vậy, ý nghĩ rằng khủng long đa số đều to lớn là sai lầm, vì hóa thạch lớn có khuynh hướng được giữ lâu hơn. Nhiều chi khủng long khá là nhỏ, vi dụ Xixianykus chỉ dài khoảng 50 xentimét (20 inch).
Kể từ khi hóa thạch khủng long lần đầu tiên được ghi nhận trong những năm đầu thế kỷ XIX, khủng long đã trở thành một phần của văn hóa đại chúng và bộ xương hoặc mô hình hóa thạch của chúng là các điểm thu hút khách tham quan ở các viện bảo tàng. Kích thước khổng lồ của một số nhóm khủng long, cũng như bản chất có vẻ huyền bí và quái vật, đã đảm bảo cho sự xuất hiện của chúng trong những quyển sách và bộ phim bán chạy nhất như Jurassic Park. Sự nhiệt tình của công chúng đã dẫn đến các tài trợ đáng kể cho nghiên cứu khủng long, và những khám phá mới thường xuyên được cập nhật bởi các phương tiện truyền thông.
Từ nguyên
[sửa | sửa mã nguồn]"Khủng long" (được người Việt dịch ra từ tiếng Trung 恐龍)[7] là từ Hán-Việt có nghĩa là "rồng lớn". Đơn vị phân loại Dinosauria được nhà cổ sinh học Sir Richard Owen định nghĩa chính thức năm 1842 là "một tộc riêng hoặc phân bộ của bò sát Sauria", một nhóm các động vật được chấp nhận ở Anh và trên thế giới lúc bấy giờ.[8][9] Tên có nguồn gốc từ Ancient Greek δεινός (deinos) 'mạnh mẽ, khủng khiếp hoặc to lớn một cách đáng sợ', và σαῦρος (sauros) 'thằn lằn hoặc bò sát'.[8][10] Mặc dù cái tên này thường được giải thích là có liên quan đến răng, móng vuốt hoặc những đặc điểm đáng sợ khác của khủng long, theo Owen nó đơn thuần chỉ để gợi lên kích thước và vẻ oai vệ của chúng.[11]
Nhiều loại bò sát tiền sử khác như dực long, thương long, thằn lằn cá, thằn lằn đầu rắn và Dimetrodon cũng được công chúng thường coi là khủng long, nhưng thực ra là không phải.[12] Dực long, là thành viên của nhánh Ornithodira, là họ hàng xa của khủng long. Các nhóm khác kể trên, cũng như khủng long và dực long, là thành viên của nhóm bò sát Sauropsida (nhánh 2 cung bên, gồm các loài bò sát và chim hiện nay), còn Dimetrodon thuộc nhóm Synapsida (nhánh một cung bên, tiến hóa thành thú có vú hiện nay).
Định nghĩa
[sửa | sửa mã nguồn]Theo phương pháp phát sinh loài, khủng long thường được định nghĩa là nhóm động vật bao hàm tổ tiên chung gần đây nhất (MRCA) của Triceratops và chim, cũng như tất cả các hậu duệ của chúng.[13] Có ý kiến cho rằng Dinosauria nên được định nghĩa trong tương quan với MRCA của Megalosaurus và Iguanodon, bởi lẽ đây là hai trong số ba chi phân loài được Richard Owen trích dẫn khi ông công nhận nhánh Dinosauria.[14] Cả hai cách định nghĩa đều dẫn đến cùng một công thức phân loại: "Dinosauria = Ornithischia + Saurischia"; trong đó bao gồm các dòng dõi khủng long thấp hơn như: Ankylosauria (khủng long bọc giáp, ăn cỏ, đi đứng bằng bốn chân), Stegosauria (khủng long phiến sừng, ăn cỏ, đi đứng bằng bốn chân), Ceratopsia (khủng long có diềm cổ, ăn cỏ, đi đứng bằng hai hoặc bốn chân), Pachycephalosauria (khủng long sọ cứng, ăn cỏ, đi đứng bằng hai chân), Ornithopoda (khủng long ăn cỏ, đi đứng bằng hai hoặc bốn chân, bao gồm khủng long "mỏ vịt"), Theropoda (hầu hết là khủng long ăn thịt, và chim), và Sauropodomorpha (khủng long ăn cỏ, đi đứng bằng bốn chân, cổ và đuôi dài).[15]
Chim hiện được công nhận là dòng dõi duy nhất còn tồn tại của khủng long. Theo phân loại truyền thống, chim được coi là một lớp động vật khác biệt đã tiến hóa từ khủng long, vốn lại một liên bộ riêng biệt. Tuy nhiên, phần lớn giới cổ sinh vật học đương đại bác bỏ cách phân loại truyền thống, mà thay vào đó phân loại theo lối phát sinh chủng loài; cách tiếp cận này đòi hỏi rằng, để một nhóm được coi là hợp lệ, tất cả hậu duệ của các loài thành viên cũng phải được quy vào nhóm. Vậy nên, chim được coi là khủng long và khủng long, do đó, chưa tuyệt chủng.[16] Chim được xếp vào phân nhóm Maniraptora, thuộc nhánh Coelurosauria, phân bộ Therapoda, bộ Saurischia.[17]
Nghiên cứu năm 2017 của Matthew Baron, David B. Norman và Paul M. Barrett đã đề xuất một thay đổi căn bản đối với hệ thống phân loài khủng long. Phân tích phát sinh chủng loại của Baron et al. đã phục hồi giả thuyết cho rằng Ornithischia gần gũi hơn với Therapoda khi so với Sauropodomorpha, trái ngược với kiểu gộp Therapoda và Sauropodomorpha cổ điển. Tuy nhiên, điều này sẽ khiến nhóm Sauropoda và hậu duệ của chúng không còn được tính là khủng long, vậy nên nhóm tác giả đã phải định nghĩa lại Dinosauria là tổ tiên chung cuối cùng của Triceratops horridus, Passer localus và Diplodocus carnegii, cũng như tất cả các hậu duệ của chúng. Họ đồng thời hồi sinh thuật ngữ Ornithoscelida để chỉ gộp Ornithischia và Theropoda.[18][19]
Mô tả chung
[sửa | sửa mã nguồn]
Sử dụng một trong những định nghĩa trên, khủng long có thể được mô tả chung là những động vật thuộc nhóm Archosauria có chân sau duỗi thẳng bên dưới cơ thể khi đứng.[20] Các nhóm bò sát tiền sử thường bị ngộ nhận là khủng long, mặc dù không chính xác, bao gồm: thằn lằn cá, thằn lằn sông, thằn lằn cổ rắn, dực long và pelycosauria (đặc biệt là chi Dimetrodon).[12] Dực long là họ hàng xa của khủng long vì chúng cùng thuộc nhánh Ornithodira. Các nhóm còn lại, giống khủng long và dực long, đều thuộc nhánh Sauropsida (gồm bò sát và chim), ngoại trừ Dimetrodon thuộc nhánh Synapsida. Tất cả các nhóm kể trên có dáng đứng chi sau khác hoàn toàn khi so với khủng long thật sự.[21]
Khủng long là động vật có xương sống phổ biến nhất trên Trái Đất vào Đại Trung Sinh, đặc biệt là trong kỷ Jura và kỷ Phấn Trắng. Các loài động vật khác bị hạn chế về kích thước và hốc sinh thái trong thời kỳ này; như động vật có vú chẳng hạn, hiếm khi đạt được kích thước của một con mèo nhà, hầu hết đều nhỏ nhắn và săn những con mồi tí hon.[22] Khủng long là một nhóm động vật rất đa dạng và phong phú; tính đến năm 2018, hơn 900 chi khủng long phi điểu đã được xác định và tổng số chi được lưu lại trong bản ghi khảo cổ được ước tính vào khoảng 1850, gần 75% trong đó chưa được khám phá, và 1124 loài tính đến năm 2016.[23][24][25] Một nghiên cứu năm 1995 suy đoán rằng, từng tồn tại 3.400 chi khủng long khác nhau trên thế giới, tính cả các loài không được lưu giữ trong bản ghi khảo cổ.[26]
Năm 2016, số lượng loài khủng long tồn tại trong Đại Trung Sinh được ước tính rơi vào khoảng 1.543–2.468.[27][28] Năm 2021, số lượng loài chim hiện đại được ước tính vào khoảng 10.806.[29] Mặc dù tổ tiên chúng đi đứng bằng hai chân, một số loài khủng long đã phát triển dáng đi đứng bằng bốn chân; một số khác, như Anchisaurus và Iguanodon chẳng hạn, còn có thể chuyển dáng đi từ hai chân sang bốn chân. Các đặc điểm kiểu hình như sừng và mào xuất hiện rất phổ biến ở khủng long, một số thậm chí sở hữu giáp xương. Dù nổi tiếng về kích thước cơ thể khổng lồ, nhiều loài khủng long Đại Trung Sinh trên thực tế chỉ to bằng con người hoặc có khi còn bé hơn. Khủng long sinh sôi nảy nở ở nhiều nơi và bằng chứng hóa thạch cho thấy phạm vi sinh sống của chúng đã lan tỏa khắp thế giới kể từ thế Jura sớm.[30] Cũng có bằng chứng cho thấy nhiều loài khủng long phi điểu, như Microraptor chẳng hạn, có khả năng bay hoặc ít nhất là liệng, và một số khác, như họ Spinosauridae chẳng hạn, có tập tính bán thủy sinh.[31]
Đặc điểm giải phẫu
[sửa | sửa mã nguồn]
j: xương gò má, po: xương sau ổ mắt, p: xương đính, sq: xương vảy, q: xương vuông, qj: xương gò má vuông

Mặc dù các khám phá gần đây đã khiến việc trình bày một bản danh sách các đặc điểm riêng biệt được thống nhất phổ quát của khủng long trở nên khó khăn hơn bao giờ hết, song hầu như tất cả các loài khủng long được phát hiện tới nay đều chia sẻ một số điểm đột biến nhất định khi so với tổ tiên chung của chúng. Một số nhóm khủng long về sau còn phát triển nhiều đặc điểm tách biệt, song vẫn bắt nguồn từ những đặc điểm tổ tiên mà được coi là điển hình của Dinosauria. Thuật ngữ để chỉ các đặc điểm như vậy là kiểu hình dẫn xuất chia sẻ (synapomorphy).[32]
Năm 2011, nhà cổ sinh vật học Sterling Nesbitt liệt kê mười hai đặc tính dẫn xuất chia sẻ chung sau đây của nhánh Archosauria:[33]
- Ở hộp sọ, phía trước cửa sổ trên thái dương (supratemporal fenestra) còn có hố trên thái dương (supratemporal fossa).
- Các mỏm lồi đốt cổ (epipophysis) hiện diện ở các đốt cổ nằm sau đốt đội (atlas) và đốt trục (axis).
- Đỉnh của chỏm delta-ngực, tức mỏm xương chĩa ra để cung cấp chỗ bám cho cơ delta-ngực (deltopectoral), có thể nằm ở bất cứ vị trí nào kể từ khoảng 30% chiều dài của xương cánh tay.
- Xương quay (radius) ngắn hơn hoặc bằng 80% xương cánh tay (humerus).
- Mẩu chuyển thứ tư (fourth trochanter), tức mỏm xương chĩa ra để cung cấp chỗ bám cho cơ đùi-đuôi (caudofemoralis), trên xương đùi là một gờ nổi.
- Mẩu chuyển thứ tư bất đối xứng, với rìa xa, thấp, tạo thành một góc dốc so với trục.
- Trên xương sên (astragalus) và xương gót (calcaneum), xương mắt cá chân trên, mặt khớp gần nhất, bề mặt kết nối trên, của xương mác (fibula) chiếm ít hơn 30% chiều rộng của phần tử.
- Các xương ngoài chẩm (exoccipital) ở phía sau hộp sọ không gặp nhau tại đường dọc đường giữa trên nền khoang nội sọ.
- Ở khung chậu, các bề mặt khớp gần gốc đốt háng (ischium) với xương chậu (ilium) và xương mu (pubis) bị ngăn cách bởi một bề mặt lõm lớn (ở phía trên của đốt háng, một phần của khớp hông mở nằm giữa các tiếp xúc với xương mu và xương chậu).
- Cạnh sống xương chày (cnemial crest) cong theo hướng trước-bên (anterolaterally).
- Dãy lồi định hướng gần theo chiều từ trục chính giữa cơ thể ra xa (proximodistally) khác biệt hiện diện ở mặt sau của cuống xương chày xa.
- Mặt khớp giữa xương mác và xương gót, tức bề mặt trên cùng của xương gót nối với xương mác, lõm xuống.
Nesbitt đồng thời bác bỏ một số kiểu hình dẫn xuất chung được đề xuất trước đó, những đặc điểm vốn có mặt ở cả họ Silesauridae, được ông phục hồi như một nhóm chị em của Dinosauria. Những đặc điểm này bao gồm một mẩu chuyển trước lớn, khối xương bàn chân II và IV có chiều dài không đều nhau, tiếp xúc giữa đốt háng và xương mu bị tiêu giảm, đỉnh cạnh sống hiện diện trên xương chày và mỏm từ dưới lên của xương sên, v.v.[13]

Ngoài ra, một loạt các đặc điểm trên bộ xương khác được chia sẻ bởi các loài khủng long. Tuy nhiên, vì chúng là điểm chung của nhiều nhóm thằn lằn chúa khác hoặc không xuất hiện trong các loài khủng long thời kỳ đầu, những đặc điểm này không được coi là dẫn xuất chung. Ví dụ, ở động vật Hai cung bên, tổ tiên khủng long có hai cặp cửa sổ thái dương (các lỗ ở hộp sọ phía sau ổ mắt) và là thành viên của nhóm Hai cung bên Thằn lằn chúa, có thêm các lỗ mở ở mõm và hàm dưới.[34] Ngoài ra, một số đặc điểm từng được cho là dẫn xuất chung hiện được biết là đã xuất hiện trước cả khủng long, hoặc vắng mặt ở những con khủng long sớm nhất và tiến hóa độc lập bởi các nhóm khủng long khác nhau. Chúng bao gồm một xương bả vai kéo dài, một xương cùng bao gồm ba hoặc nhiều đốt sống hợp nhất (ba được tìm thấy trong một số thằn lằn chúa khác, nhưng chỉ có hai được tìm thấy ở Herrerasaurus);[13] và một ổ cối mở, hoặc ổ cắm hông, có một lỗ ở giữa ở bề mặt bên trong của nó (đóng vào ở Saturnalia).[35][36] Một khó khăn khác trong việc xác định các đặc điểm chung của khủng long là khủng long sơ khai và các loài săn mồi khác từ thế Đệ Tam muộn thường được biết đến rất ít và giống nhau theo nhiều cách; những con vật này đôi khi bị xác định sai trong nhiều tài liệu.[37]
Khủng long đứng với chân sau dựng đứng theo cách tương tự như hầu hết các động vật có vú hiện đại, khác biệt với hầu hết các loài bò sát khác có tư thế chân nằm ngang.[38] Tư thế này xuất hiện là do sự phát triển của một ngách ở mặt bên trong khung chậu (thường là một ổ mở) và một đầu khác biệt hướng vào bên trong nằm trên xương đùi.[39] Tư thế duỗi thẳng cho phép những con khủng long nguyên thủy dễ hô hấp hơn trong khi đang di chuyển, tăng sức chịu đựng và mức độ hoạt động vượt qua những loài bò sát xoải chân.[40] Tay chân duỗi thẳng có lẽ đã hỗ trợ sự tiến hóa của kích thước cơ thể lớn bằng cách giảm lực uốn cong lên các chi.[41] Một số thằn lằn chúa không phải khủng long, bao gồm rauisuchian, cũng có chân tay duỗi thẳng nhưng đạt được điều này nhờ cấu hình "trụ cột" của khớp hông, trong đó thay vì xương đùi chèn vào ổ cắm trên hông, xương chậu được xoay lại để tạo thành một kệ nhô ra.[41]
Lịch trình tiến hóa
[sửa | sửa mã nguồn]Nguồn gốc và sự tiến hóa sớm
[sửa | sửa mã nguồn]Khủng long phân nhánh từ tổ tiên của chúng, một loài bò sát thuộc nhóm Archosauria, vào khoảng 230 triệu năm trước, trong khoảng Thế Tam Điệp giữa đến muộn, gần 20 triệu năm sau sự kiện tuyệt chủng Permi–Trias quét sạch khoảng 96% dạng sống dưới nước và 70% dạng sống có xương trên cạn.[42][43] Độ tuổi của các thành hệ chứa hóa thạch của chi Eoraptor rơi vào khoảng 231.4 triệu năm. Các nhà cổ sinh vật học cho rằng Eoraptor sở hữu nhiều đặc điểm của tổ tiên chung của tất cả các loài khủng long;[44] nếu đây là sự thật thì nó chỉ ra rằng những con khủng long đầu tiên đều là những săn mồi cỡ nhỏ đi bằng hai chân.[45] Việc phát hiện ra các ornithodiran nguyên thủy giống khủng long như Marasuchus và Lagerpeton thuộc tầng đá có tuổi Tam Điệp trung ở Argentina chứng minh cho quan điểm này; phân tích những hóa thạch được phục dựng lại cho thấy rằng những động vật này là các loài săn mồi cỡ nhỏ đi bằng hai chân. Tuy vậy, khủng long có thể đã xuất hiện sớm hơn nữa, vào khoảng 243 triệu năm trước, dựa trên các mảnh hóa thạch còn sót lại của chi Nyasasaurus, tuy vậy những mảnh xương này đã bị biến dạng quá nhiều, khiến cho việc xác định nó có quan hệ thế nào với khủng long thành ra không chắc chắn.[46]
Khi khủng long lần đầu tiên xuất hiện, chúng chưa phải là loài động vật có xương sống chiếm ưu thế. Các lục địa bấy giờ bị chiếm lĩnh bởi nhiều loài archosauria và thuộc Bộ Cung thú khác như aetosauria, cynodontia, ornithosuchidae, rauisuchia, và rhynchosauria. Hầu hết các nhóm này đã tuyệt chủng trong kỷ Tam Điệp thuộc một trong 2 làn sóng tuyệt chủng. Làn sóng đầu tiên, vào khoảng ranh giới giữa các tầng động vật Carnian và Norian (khoảng 215 triệu năm trước), nhánh gốc của archosauromorpha bao gồm các protorosaur bị tuyệt chủng. Tiếp theo đó là giai đoạn tuyệt chủng Trias-Jura (khoảng 200 triệu năm trước), hầu hết các nhóm thằn lằn chúa thời kỳ đầu như aetosauria, ornithosuchidae, phytosauria, và rauisuchia bị tuyệt chủng. Sự mất mát này để lại một hệ động vật trên cạn gồm crocodylomorpha (nhánh chứa cá sấu sau này), khủng long, thú, pterosauria, và rùa.[13] Dòng dõi mới của khủng long ban đầu đa dạng hóa qua các tầng động vật Carnian và Norian trong kỷ Trias bằng cách chiếm các hốc sinh thái của các nhóm đã tuyệt chủng. ...
Tiến hóa và cổ địa sinh học
[sửa | sửa mã nguồn]Sự tiến hóa của khủng long hậu Tam Điệp đi kèm theo những thay đổi của thảm thực vật và vị trí của các lục địa. Vào kỷ Tam Điệp muộn và kỷ Jura sớm, các khối đất liền ghép với nhau tạo thành một lục địa Pangea duy nhất, và hệ động vật của thế giới chủ yếu bao gồm các loài ăn thịt coelophysoidea và các loài ăn cỏ sauropodomorph sớm.[47] Thực vật hạt trần (đặc biệt là cây lá kim), một nguồn thức ăn tiềm năng, phát triển trong thế Tam Điệp muộn. Các sauropodomorph nguyên thủy không có cơ chế tinh vi để nghiền thức ăn trong miệng, và do đó phải sử dụng phương pháp khác để băm nhỏ thức ăn trong hệ tiêu hóa.[48] Sự đồng nhất tổng quát của các loài khủng long vẫn tiếp tục trong kỷ Jura giữa và muộn, các địa phương hầu hết đều có động vật ăn thịt lớn bao gồm Ceratosauria, đại long và Carnosauria và các loài khủng long hông chim, khủng long phiến sừng và khủng long chân thằn lằn lớn ăn thực vật. Ví dụ bao gồm thành hệ Morrison tại Bắc Mỹ và các thềm Tendaguru tại Tanzania. Khủng long ở Trung Quốc có một số khác biệt, với các loài khủng long chân thú sinraptorid chuyên biệt và các loài khủng long cổ dài khác thường như Mamenchisaurus.[47] Khủng long bọc giáp và khủng long chân chim cũng trở nên phổ biến hơn, trong khi các chuẩn bản long (tổ tiên khủng long chân thằn lằn) tuyệt chủng. Cây lá kim và pteridophyte là những loài thực vật phổ biến nhất. Khủng long chân thằn lằn, giống như chuẩn bản long trước đây, không nghiền lá cây cỏ bằng miệng. Các loài khủng long hông chim bấy giờ đã phát triển các cách thức khác nhau để xử lý thức ăn trong miệng, bao gồm các cơ quan giống như má để giữ thức ăn trong mồm và các chuyển động hàm thích hợp để nghiền thức ăn.[48] Một sự kiện tiến hóa đáng chú ý khác trong kỷ Jura là sự xuất hiện của những loài chim thực sự, có nguồn gốc từ nhánh khủng long đuôi rỗng maniraptora.

Vào kỷ Phấn trắng sớm, Pangea bắt đầu phân tách ra. Khủng long theo đó mà trở nên đa dạng hơn bao giờ hết. Phần đầu của thời đại này đã chứng kiến sự mở rộng của các loài khủng long vận giáp, iguanodontia và brachiosaurid tại châu Âu, Bắc Mỹ và phía bắc châu Phi. Những nhóm này sau đó bị thay thế ở châu Phi bằng các khủng long chân thú khác như các đại long xương gai và dị long răng cá mập lớn, và các khủng long chân thằn lằn như họ Rebbachisauridae và nhóm Titanosauria, cũng được tìm thấy ở Nam Mỹ. Ở châu Á, khủng long đuôi rỗng maniraptor, các khủng long chạy nhanh, các điểu long răng khía và các oviraptorosauria trở nên phổ biến, và các khủng long vận giáp và khủng long mặt sừng sớm như Psittacosaurus trở thành các động vật ăn cỏ chủ chốt. Trong khi đó, Úc là nơi cư trú của một số loài khủng long vận giáp nguyên thủy, họ Hypsilophodont và các iguanodontia.[47] Các khủng long phiến sừng dường như đã tuyệt chủng vào một thời điểm nào đó trong kỷ Phấn trắng sớm hoặc Phấn trắng muộn sớm. Một thay đổi lớn trong kỷ Phấn trắng sớm, sẽ được khuếch đại vào cuối kỷ Phấn trắng, là sự tiến hóa của thực vật có hoa. Đồng thời, một số nhóm khủng long ăn cỏ đã phát triển những cách thức tinh vi hơn để tiêu hóa thức ăn. Những khủng long đầu giáp đã phát triển một phương pháp cắt lát với những chiếc răng xếp chồng lên nhau trong một khối răng, và những loài iguanodontia phát triển một phương pháp mài bằng khối răng, được nâng cấp ở các khủng long mỏ vịt.[48] Một số loài khủng long chân thằn lằn cũng tiến hóa khối răng, được minh họa tốt nhất ở loài Nigersaurus.[49]

Có ba hệ động vật khủng long chính vào cuối kỷ Phấn trắng. Ở các lục địa phía bắc của Bắc Mỹ và Châu Á, các khủng long chân thú chủ chốt là các bạo long chuẩn và nhiều loại maniraptora nhỏ khác nhau, với các loài động vật ăn cỏ chủ yếu là khủng long hông chim gồm mỏ vịt, mặt sừng, vận giáp và đầu dày. Ở các lục địa phía nam từng tạo nên lục địa Gondwana đang bị chia tách, các abelisaurid là loài khủng long chân thú phổ biến và các titanosauria là động vật ăn cỏ phổ biến. Cuối cùng, ở châu Âu, khủng long chạy nhanh, rhabdodontid, giáp long xương kết, và loài titanosauria là phổ biến.[47] Thực vật có hoa phát xạ tiến hóa cao,[48] với những đám cỏ đầu tiên xuất hiện vào cuối kỷ Phấn trắng.[50] Các loài hadrosaurid ăn nghiền và ceratopsia ăn cắt thực vật trở nên vô cùng đa dạng trên khắp Bắc Mỹ và Châu Á. Khủng long chân thú cũng phát xạ tiến hóa thành các loài ăn cỏ hoặc ăn tạp, với các loài therizinosauria và ornithomimosauria trở nên phổ biến hơn.[48]
Sự kiện tuyệt chủng kỷ Phấn Trắng - Cổ Cận, xảy ra vào khoảng 66 triệu năm trước, giết chết tất cả các nhóm khủng long ngoại trừ các loài chim neornithine. Một số nhóm Hai cung thú khác, chẳng hạn như cá sấu, sebecosuchia, rùa, thằn lằn, rắn, sphenodontia, và choristodera, cũng sống sót sau sự kiện này.[51]
Các dòng dõi còn sót lại của các loài chim neornithine, bao gồm tổ tiên của các loài ratite (các loài chim lớn không bay), điểu cầm hiện đại, và một loạt các loài chim nước, đa dạng hóa nhanh chóng vào đầu kỷ Cổ Cận, chiếm các hốc sinh thái bị bỏ trống bởi sự tuyệt chủng của loài khủng long đại Trung Sinh ví dụ như các enantiornithine sống trên cây, hesperornithes thủy sinh, và thậm chí thay thế cả các loài khủng long chân thú trên cạn lớn (dưới dạng Gastornis, eogruiid, Bathornithid, ratite, geranoidid, mihirung và "chim khủng bố"). Người ta thường viện dẫn rằng các động vật có vú cạnh tranh với các neornithine để thống trị các hốc sinh thái trên cạn nhưng nhiều nhóm trong số này cùng tồn tại ôn hòa với các động vật có vú phong phú trong hầu hết đại Tân Sinh.[52] Những con chim khủng bố và Bathornithid chiếm các hốc ăn thịt cùng với các động vật có vú ăn thịt,[53][54] và rattie vẫn khá thành công trong vai trò là động vật ăn cỏ cỡ trung bình; eogruiid tương tự kéo dài từ thế Thủy Tân đến thế Thượng Tân, chỉ bị tuyệt chủng rất gần đây sau hơn 20 triệu năm cùng tồn tại với nhiều nhóm động vật có vú.[55]
Phân loại
[sửa | sửa mã nguồn]Khủng long là archosauria (thằn lằn chúa), nhóm bao gồm cá sấu hiện đại. Trong nhóm archosauria, khủng long được phân biệt chủ yếu bởi dáng đi của chúng. Chân khủng long ở ngay dưới cơ thể chúng, trong khi chân thằn lằn và cá sấu chỉa ra sang hai bên.[32]
Nói chung, khủng long là một nhánh được chia thành hai phần Saurischia và Ornithischia. Saurischia gồm các loài khủng long có chung tổ tiên gần với chim hơn Ornithischia, trong khi Ornithischia có chung tổ tiên với Triceratops hơn Saurischia. Về mặt giải phẫu, hai nhóm này có thể được phân biệt rõ rệt nhất bởi cấu trúc xương chậu của chúng. Saurischia thời kỳ đầu - "hông thằn lằn", từ tiếng Hy Lạp sauros (σαῦρος) có nghĩa là "thằn lằn" và ischion (ἰσχίον) có nghĩa là "khớp hông" - giữ lại cấu trúc hông tổ tiên của chúng, với xương mu hướng về phía trước.[39] Hình thức cơ bản này đã được sửa đổi bằng cách xoay các xương mu về phía sau ở các mức độ khác nhau trong một số nhóm (Herrerasaurus,[56] Therizinosauroid, và chim[17]). Saurischia bao gồm các loài khủng long chân thú (đi bằng hai chân và với nhiều chế độ ăn khác nhau) và khủng long dạng chân thằn lằn (động vật ăn cỏ cổ dài bao gồm các nhóm bốn chân tiên tiến).[31][57]
Ngược lại, Ornithischia - "hông chim", từ tiếng Hy Lạp ornitheios (ὀρνίθειος) nghĩa là "thuộc về chim" và ischion (ἰσχίον) có nghĩa là "khớp hông" - có một cấu trúc khung chậu trông giống ở chim: xương mu định hướng chĩa ra sau. Không giống như các loài chim, Ornithischia cũng thường có một mỏm xương hướng về phía trước. Ornithischia bao gồm nhiều loài chủ yếu là động vật ăn cỏ. (chú thích: thuật ngữ "hông thằn lằn" và "hông chim" là những cách hiểu sai - loài chim tiến hóa từ khủng long với "hông thằn lằn".)[32]
-
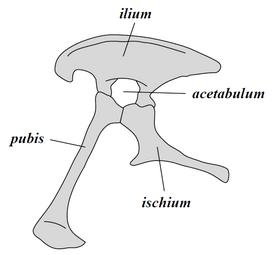 khung chậu của Saurischia (bên trái)
khung chậu của Saurischia (bên trái) -
 cấu trúc khung chậu của Tyrannosaurus (cho thấy cấu trúc hông thằn lằn – bên trái)
cấu trúc khung chậu của Tyrannosaurus (cho thấy cấu trúc hông thằn lằn – bên trái) -
 khung chậu Ornithischia (bên trái)
khung chậu Ornithischia (bên trái) -
 cấu trúc khung chậu của Edmontosaurus (cho thấy cấu trúc hông chim – bên trái)
cấu trúc khung chậu của Edmontosaurus (cho thấy cấu trúc hông chim – bên trái)




Phân loại
[sửa | sửa mã nguồn]Phân loại rút gọn dưới đây của các nhóm khủng long dựa trên quan hệ tiến hóa, sắp xếp dựa trên danh sách khủng long Đại Trung Sinh của Holtz (2008). † biểu thị cho nhóm không còn thành viên sinh tồn.
- Dinosauria
- Saurischia ("hông thằn lằn"; bao gồm Theropoda và Sauropodomorpha)
- †Herrerasauria (khủng long ăn thịt đi hai chân sớm)
- Theropoda (tất cả đi hai chân; hầu hết ăn thịt)

- †Coelophysoidea (nhỏ, khủng long chân thú sớm, gồm Coelophysis và họ hàng)
- †Dilophosauridae (có mào, khủng long chân thú ăn thịt sớm)
- †Ceratosauria (có sừng, thú ăn thịt hàng đầu của các lục địa phía Nam trong kỷ Phấn Trắng)
- Tetanurae ("đuôi cứng"; bao gồm hầu hết khủng long chân thú)
- †Megalosauroidea (nhóm khủng long khổng lồ ăn thịt sớm)
- †Carnosauria (Allosaurus và họ hàng)
- Coelurosauria (nhóm khủng long có lông vũ, kích thước đa dạng và chiếm nhiều hốc sinh thái)[58]
- †Compsognathidae (coelurosauria sớm phổ biến với chi trước ngắn đi)
- †Tyrannosauridae (Tyrannosaurus và họ hàng, chi trước ngắn)
- †Ornithomimosauria ("giống như đà điểu"; hầu hết không có răng, có thể ăn thịt hoặc cỏ)
- †Alvarezsauroidea (ăn côn trùng, chi trước có vuốt lớn)
- Maniraptora ("bắt bằng tay"; có tay với ngón tay gầy và dài)
- †Therizinosauria (ăn cỏ, sở hữu các móng vuốt lớn và cái đầu nhỏ)
- †Oviraptorosauria (hầu hết không có răng, lối sống và thức ăn chưa rõ ràng)
- †Archaeopterygidae (khủng long có cánh hoặc chim nguyên thủy)
- †Deinonychosauria (cỡ nhỏ đến trung bình, giống chim, ăn thịt và có vuốt to ở chân)
- Avialae (chim hiện đại và họ hàng đã tuyệt chủng)
- †Scansoriopterygidae (avialan sơ khai cỡ nhỏ, có ba móng vuốt ở chi trước)
- †Omnivoropterygidae (avialan sơ khai cỡ lớn, đuôi nhỏ)
- †Confuciusornithidae (avialan cỡ nhỏ không có răng)
- †Enantiornithes (avialan sơ khai sống trên cây, biết bay)
- Euornithes (chim biết bay tân tiến)
- †Yanornithiformes (chim kỷ Phấn Trắng ở Trung Quốc có răng)
- †Hesperornithes (chim lặn)
- Aves (chim hiện đại)

- †Sauropodomorpha (ăn thực vật với đầu nhỏ, cổ và đuôi dài)
- †Guaibasauridae (nhỏ, sơ khai, ăn tạp)
- †Plateosauridae (sơ khai, đi bằng hai chân)
- †Riojasauridae (sơ khai, nhỏ)
- †Massospondylidae (sơ khai, nhỏ)
- †Sauropoda (khủng long bốn chân kích cỡ khổng lồ và rất nặng)
- †Vulcanodontidae (khủng long chân thằn lằn sơ khai với bôn chân hình cột)
- †Eusauropoda ("khủng long chân thằn lằn thật sự")
- †Cetiosauridae ("thằn lằn cá voi")
- †Turiasauria (nhóm chân thằn lằn từ Châu Âu vào kỷ Tam Điệp và Phấn Trắng)
- †Neosauropoda ("khủng long chân thằn lằn mới")
- †Diplodocoidea (đuôi và cổ cực dài, răng rất hẹp và giống bút chì)
- †Macronaria (hộp sọ vuông vắn, răng hình cái thìa hoặc bút chì)
- †Brachiosauridae (cổ dài, chi trước cao hơn)
- †Titanosauria (đa dạng; nặng cân, hông rộng; phổ biến ở các lục địa phía Nam vào kỷ Phấn Trắng)

- †Ornithischia ("hông chim"; ăn thực vật)
- †Heterodontosauridae (chân chim nguyên thủy nhỏ ăn tạp có răng giống răng nanh)
- †Thyreophora (bốn chân, mang giáp tự vệ)
- †Ankylosauria (có giáp, vài loài có đuôi trùy)
- †Stegosauria (có gai hoặc tấm sừng trên lưng)
- †Neornithischia ("hông chim mới")
- †Ornithopoda (bốn hoặc hai chân, ăn thực vật; phát triển cách nghiền thức ăn sử dụng sự dẻo dai của hộp sọ và nhiều răng)
- †Marginocephalia (phân biệt nhờ đặc điểm của đầu)
- †Pachycephalosauria (hai chân, đầu dày hoặc có bướu trên đầu)
- †Ceratopsia (bốn chân, đầu có diềm; nhiều loài có sừng)
Sinh học
[sửa | sửa mã nguồn]Kích thước
[sửa | sửa mã nguồn]Nhóm Sauropoda có các loài khủng long lớn và nặng nhất. Trong hầu hết thời kỳ khủng long, sauropoda nhỏ nhất vẫn lớn hơn hầu hết những loài động vật khác trong môi trường của nó, và sauropoda lớn nhất theo thứ lượng nặng hơn bất cứ động vật nào từng bước đi trên mặt đất. Các loài động vật có vú không lồ tiền sử như Paraceratherium vẫn được xem là lùn với saropoda, chỉ có cá voi mới bằng hoặc vượt hơn saropoda.[59] Có nhiều lợi thế sauropoda nhận được từ kích thước lớn, bao gồm được bảo vệ khỏi khủng long ăn thịt, giảm sử dụng nặng lượng, và sống lâu, nhưng có thể lợi thế quan trọng nhất là chế độ ăn uống. Động vật lớn có hiệu quả tiêu hóa cao hơn động vật nhỏ, vì thức ăn có nhiều thời gian nằm trong hệ tiêu hóa của nó hơn. Điều này cũng cho phép chúng sống sót với thức ăn có giá trị dinh dưỡng thấp.[60]
Lớn nhất và nhỏ nhất
[sửa | sửa mã nguồn]Các nhà khoa học có thể sẽ không bao giờ tìm thấy khủng long to và nhỏ nhất. Bởi vì chỉ có một số nhỏ khủng long để lại hóa thạch sau khi chết, và hầu hết số hóa thạch này vẫn chưa được tìm thấy mà vẫn còn bị chôn vùi trong lòng đất. Rất ít mẫu vật được thu hồi là những bộ xương hoàn chỉnh, và dấu vết về da và mô mềm rất hiếm. Phục dựng một bộ xương hoàn chỉnh theo so sánh kích thước và hình thái xương của các loài tương tự nhau là một phương pháp phổ biến nhưng không đủ chính xác, và việc phục nguyên các bắp thịt, cơ quan chỉ là sự phỏng đoán dựa theo một số nguyên tắc sinh học[61]

Loài khủng long cao nhất và nặng nhất được biết đến từ một bộ xương bảo quản tốt là Giraffatitan brancai (trước được xem là một loài Brachiosaurus). Hóa thạch của nó được phát hiện ở Tanzania giữa năm 1907 và 1912. Xương của nhiều cá thể tương tự về kích thước đã được ráp vào nhau thành một bộ xương hoàn chỉnh nay được trưng bày tại bảo tàng für Naturkunde Berlin;[62] bộ xương này cao 12 mét (39 ft) và dài 22,5 mét (74 ft), và có thể thuộc về một động vật nặng khoảng 30000 đến 60000 kg (70000 đến 130000 lb). Con khủng long có hóa thạch hoàn chỉnh dài nhất (27 mét, 89 ft) là Diplodocus, phát hiện ở Wyoming ở Mỹ và được trưng bày tại bảo tàng lịch sử tự nhiên Carnegie của Pittsburgh năm 1907.[63]
Có nhiều loài khủng long lớn hơn, nhưng hiểu biết về chúng hoàn toàn nhờ các mẫu hóa thạch rời rạc. Hầu hết được thu thập từ 1970 hay sau đó, bao gồm cả Argentinosaurus, có thể nặng từ 80000 tới 100000 kg (90 tới 110 tấn ngắn); vài cá thể Diplodocus hallorum (trước đây Seismosaurus) dài tới 33,5 mét (110 ft)[60] và Supersaurus dài 33 mét (108 ft);[64] và cao nhất là Sauroposeidon cao 18 mét (59 ft), chạm vào cửa sổ toà nhà sáu tầng. Loài nặng nhất và dài nhất trong số chúng có thể là Amphicoelias fragillimus, được biết duy nhất từ một đốt sống nay đã thất lạc năm 1878. Suy ra từ hình minh họa đốt sống này, con vật này có thể dài 58 mét (190 ft) và nặng hơn 120000 kg (260000 lb).[60] Khủng long ăn thịt lớn nhất là Spinosaurus, chiều dài đạt đến 16-18 mét (52 to 60 ft), và nặng 8150 kg (18000 lb).[65] Vài loài ăn thịt lớn khác là Giganotosaurus, Carcharodontosaurus và Tyrannosaurus.[66]
Trong các loài khủng long phi chim (Avialae), loài nhỏ nhất chỉ có kích thước bằng bồ câu.[67] Không ngạc nhiên, chúng là những theropoda họ hàng gần nhất với chim. Ví dụ, Anchiornis huxleyi, có bộ xương hoàn chỉnh dài 35 cm(1.1 ft).[67][68] A. huxleyi là loài khủng long phi chim nhỏ nhất được mô tả từ cá thể trưởng thành, với cân nặng 110 gram.[68] Khủng long ăn cỏ nhỏ nhất là Microceratus và Wannanosaurus, dài khoảng 60 cm (2,0 ft).[69]
Hành vi
[sửa | sửa mã nguồn]
Nhiều loài chim hiện đại có tính xã hội cao, và thường sống thành bầy. Có đồng thuận chung rằng nhiều hành vi thông thường ở chim, cũng như ở cá sấu (họ hàng còn sinh tồn gần nhất của chim), cũng xuất hiện ở khủng long. Giải thích hành vi ở các loài hóa thạch chủ yếu dựa trên tư thế bộ xương và môi trường sống, máy tính mô hình hóa của cơ sinh học khủng long, và so sánh với động vật hiện đại có chung hốc sinh thái.[70]
Bằng chứng đầu tiên cho thấy sống theo bầy là một hành vi thông thường ở khủng long là vào năm 1878 khi phát hiện 31 Iguanodon bernissartensis được cho là chết cùng nhau tại Bernissart, Bỉ, sau khi tất cả rơi vào một hố sụt sâu đầy nước và chết đuối.[71] Các vùng hóa thạch như thế sau đó được phát hiện thêm. Dấu chân của hàng trăm hay hàng ngàn cá thể khủng long ăn cỏ được cho là khủng long mỏ vịt (hadrosauridae) cho thấy chúng có thể di chuyển theo đàn lớn, giống Bò rừng bizon Bắc Mỹ hay Linh dương nhảy. Dấu chân sauropoda cho thấy chúng di chuyển thành đàn nhiều loài khác nhau, ít nhất là tại Oxfordshire, Anh,[72] mặc dù không có bằng chứng chính xác về cấu trúc đàn.[73] Có nhiều loại khủng long lớn chậm, bao gồm nhiều theropoda, sauropoda, ankylosauria, ornithopoda, và ceratopsia, tạo thành một đàn các cá thể non. Ví dụ như ở vùng Nội Mông có hơn 20 bộ hóa thạch Sinornithomimus, từ một tới bảy tuổi. Sự tập hợp này được giải thích là một nhóm xã hội bị kẹt trong bùn.[74] Điều này cũng khiến therapoda trở thành loài săn cùng nhau để giết các con mồi lớn.[75][76] Tuy nhiên, cách sống này không phổ biến với chim hiện đại, cá sấu, bò sát khác.

Mào và diềm cổ ở vài nhóm khủng long, như marginocephalia, theropoda và lambeosaurinae, có thể quá mỏng manh để tự vệ, và vì vậy chúng được dùng để biểu thị sự hung hăng hay tình dục, mặc dù sự giao phối và lãnh thổ của khủng long được biết đến rất ít.
Một hóa thạch quan trong được phát hiện tại Sa mạc Gobi năm 1971. Một con Velociraptor tấn công Protoceratops,[77] có bằng chứng cho thấy mỗi con thực sự tấn công con còn lại.[78] Vài loài khủng long ăn thịt đồng loại được thừa nhận, bởi các dấu răng Majungasaurus trên xương của Majungasaurus khác tại Madagascar năm 2003.[79]
So sánh ổ mắt khủng long với chim và bò sát được dùng để phỏng đoán các hành vi hàng ngày của khủng long. Mặc dù có gợi ý rằng khủng long hoạt động suốt ban ngày, các loài khủng long ăn thịt nhỏ như dromaeosauridae, Juravenator, và Megapnosaurus được cho là sống về đêm. Các loài ăn cỏ và ăn tạp vừa hay lớn như ceratopsia, Sauropodomorpha, Hadrosauridae, Ornithomimosauria hoạt động suốt một ngày, với các khoảng nghỉ ngắn, mặc dù Ornithomimosauria Agilisaurus sống ban ngày.[80]
Vài ornithischia, như Oryctodromeus có một phần cuộc sống đào bới trong hang.[81] Nhiều loài chim sống trên cây, điều này cũng đúng với chim Đại Trung Sinh, đặc biệt enantiornithes.[82] Vài loài khủng long giống chim cũng sống tốt trên cây (gồm cả Microraptor[83]) hầu hết khủng long phi chim sống trên mặt đất.
Sinh sản
[sửa | sửa mã nguồn]Khủng long đẻ trứng có màng ối với vỏ cứng được cấu tạo chủ yếu bởi calci cacbonat.[84] Hầu hết xây tổ rất phức tạp, có thể dạng cúp, vòm, đĩa, ụ, và cả đào hang.[85] Vài loài chim hiện đại không xây tổ; chim uria thông thường đẻ trứng trên vách đá, và chim cánh cụt hoàng đế đực kẹp trứng giữa hai chân. Chim tiền sử và nhiều loài khủng long phi chim đẻ trứng vào tổ chung, và thường thì con đực ấm trứng. Chim hiện đại chỉ có một vòi trứng và chỉ đẻ một trứng/lần, chim nguyên thủy và khủng long có hai vòi trứng, giống cá sấu. Vì chi khủng long, như Troodon đẻ hai trứng mỗi một hay hai ngày, và làm trứng nở đồng thời bằng cách không ấp trứng tới khi tất cả trứng được đẻ.[86]
Sự tuyệt chủng
[sửa | sửa mã nguồn]Việc khám phá rằng chim là một loại khủng long cho thấy khủng long vẫn chưa tuyệt chủng như thường được nói.[87] Tuy nhiên, tất cả khủng long phi chim và nhiều nhóm chim đã tuyệt chủng cách đây khoảng 66 triệu năm. Nhiều nhóm động vật khác cũng tuyệt chủng vào thời gian này, gồm cúc đá (thân mềm giống ốc anh vũ), mosasauridae, plesiosauria, pterosauria, và nhiều nhóm động vật có vú.[30] Côn trùng hầu như không chịu sự ảnh hưởng nào, chúng trở thành thức ăn cho các loài còn sống sót. Sự kiện tuyệt chủng này được gọi là sự kiện tuyệt chủng kỷ Creta-Paleogen. Bản chất của sự kiện gây tuyệt chủng hàng loạt này đã được nghiên cứu từ những năm 1970; tới nay, nhiều giả thuyết được các nhà cổ sinh vật học hỗ trợ. Mặc dù hầu hết hưởng ứng rằng sự kiện va chạm tiểu hành tinh (hay thiên thạch) là nguyên nhân chính, vài nhà khoa học chỉ ra các nguyên nhân khác, hay ủng hộ ý kiến rằng có nhiều yếu tố góp phần vào sự biến mất đột ngột của khủng long.[88][89][90]
Vào Đại Trung Sinh, không có mũ băng địa cực, và mực nước biển được cho là cao hơn ngày nay từ 100 tới 250 mét (300 tới 800 ft). Nhiệt độ hành tinh cũng đồng bộ hơn, nhiệt độ tại xích đạo chỉ cách tại địa cực 25 °C (45 °F). Trung bình, nhiệt độ không khí cao hơn; tại địa cực, nhiệt độ cao hơn ngày hôm nay 50 °C (90 °F).[91][92]
Cấu tạo khí quyển Đại Trung Sinh là vấn đề tranh luận. Vài lý thuyết cho rằng nồng độ oxy cao hơn ngày nay, số khác cho rằng sự thích nghi sinh học ở chim và khủng long cho thấy hệ thống hô hấp phát triển xa hơn cần thiết nếu nồng độ oxy đạt mức cao.[93] Vào cuối kỷ Phấn Trắng, môi trường là thay đổi đáng kể. Hoạt động núi lửa giảm, dẫn đến xu hướng cacbon dioxide giảm (do núi lửa phun nhiều chất này). Nồng độ oxy trong khí quyển cũng bắt đầu dao động và cuối cùng giảm xuống. Một số nhà khoa học đưa ra giả thuyết rằng biến đổi khí hậu, kết hợp với nồng độ oxy thấp, có thể trực tiếp dẫn đến sự tuyệt chủng của nhiều loài.[94]
Sự va chạm của tiểu hành tinh
[sửa | sửa mã nguồn]
Lý thuyết va chạm tiểu hành tinh, nhận được sự chú ý rộng vào năm 1980 bởi Walter Alvarez cùng với cha là Luis Walter Alvarez và đồng nghiệp, cho rằng một tiểu hành tinh đã va vào Trái Đất vào khoảng 66 triệu năm về trước. Alvarez phát hiện các lớp trầm tích phân bố trên khắp thế giới có chứa hàm lượng iridi cao gấp nhiều lần, là bằng chứng cho vụ va chạm.[95] Tiểu hành tinh (hay thiên thạch) được cho là rộng 5 tới 15 km (3 tới 9 mi) đã đâm vào bán đảo Yucatan ở México, tạo nên hố Chicxulub 180 km (110 mi) và gây nên sự kiện tuyệt chủng.[96][97] Các nhà khoa học không chắc rằng khủng long đang phát triển hay suy tàn trước vụ va chạm.
Phun trào núi lửa tại trap Deccan
[sửa | sửa mã nguồn]Trước năm 2000, các luận cứ cho rằng các dòng lũ bazan tại trap Deccan đã gây ra sự tuyệt chủng thường gắn liền với quan điểm cho rằng quá trình khủng long tuyệt chủng diễn ra từ từ, do các dòng lũ bazan được cho là bắt đầu diễn ra trong khoảng 68 Ma và kéo dài hơn 2 triệu năm. Bằng chứng gần đây nhất cho thấy các bẫy này đã phun trào hơn 800.000 năm kéo qua ranh giới K-T, và do đó có thể nó đóng vai trò trong sự tuyệt chủng và sự hồi phục sinh học chậm sau đó.[98]
Các trap Deccan có thể là nguyên nhân gây tuyệt chủng theo một số cơ chế như sự giải phóng bụi và các sol khí sulfuric vào không khí làm ngăn chặn ánh sáng Mặt Trời và giảm quá trình quang hợp của thực vật. Ngoài ra, hoạt động núi lửa trap Deccan có thể sinh ra một lượng cacbon dioxide lớn gây ra hiệu ứng nhà kính khi bụi và sol khí chiếm toàn bộ khí quyển.[99]
Trong những năm khi mà giả thuyết trap Deccan còn gắn với sự tuyệt chủng từ từ, Luis Walter Alvarez (1911 - 1988) đã cho rằng các nhà cổ sinh vật học bị lầm lạc do thiếu hụt dữ liệu. Trong khi khẳng định của ông ban đầu không được ủng hộ, nhưng các nghiên cứu thực địa sau đó về các lớp đáy hóa thạch lại nghiêng về phía tuyên bố của ông. Cuối cùng thì hầu hết các nhà cổ sinh vật học bắt đầu chấp nhận ý tưởng rằng sự tuyệt chủng hàng loạt vào cuối kỷ Creta chủ yếu hay một phần là do sự va chạm của tiểu hành tinh vào Trái Đất. Tuy nhiên, ngay cả Walter Alvarez cũng đã thừa nhận rằng có các thay đổi lớn khác trên Trái Đất đã xảy ra ngay trước sự va chạm, như mực nước biển tụt xuống và các hoạt động phun trào núi lửa diễn ra mạnh mẽ đã tạo ra các trap Deccan ở Ấn Độ, và các thay đổi này có thể đã góp phần tạo ra sự tuyệt chủng.[100]
Khủng long sống sót qua vụ tuyệt chủng
[sửa | sửa mã nguồn]Hóa thạch khủng long phi chim đôi khi được tìm thấy qua ranh giới Creta–Paleogen. Năm 2001, nhà cổ sinh vật học Zielinski và Budahn thông báo đã phát hiện một hóa thạch xương chân hadrosauridae ở lưu vực San Juan, New Mexico, và mô tả nó như một bằng chứng của khủng long thế Paleocen. Hóa thạch được cho là vào đầu kỷ Đệ Tam, khoảng 64.5 triệu năm trước. Nếu nó không bị hoạt động thời tiết làm thay đổi địa tầng, nó được xem là bằng chứng rằng một vài quần thể khủng long đã sống sót đến kỷ Đệ Tam.[101] Chứng cứ khác bao gồm hóa thạch được tìm thấy trong thành hệ Hell Creek khoảng (40000 năm sau ranh giới Creta–Paleogen). Các báo cáo tương tự cũng đến từ các phần khác của thế giới, gồm cả Trung Quốc.[102]
Ghi chú
[sửa | sửa mã nguồn]- ^ Các nghiên cứu gần đây xem khủng long không phải là động vật máu lạnh, dù cho chúng là bò sát; xem § Đặc điểm sinh lý
Tham khảo
[sửa | sửa mã nguồn]- ^ Matthew G. Baron; Megan E. Williams (2018). "A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution". Acta Palaeontologica Polonica. 63. doi:10.4202/app.00372.2017.
- ^ Andrea Cau (2018). "The assembly of the avian body plan: a 160-million-year long process" (PDF). Bollettino della Società Paleontologica Italiana. 57 (1): 1–25. doi:10.4435/BSPI.2018.01.
- ^ Ferigolo, Jorge; Langer, Max C. (1 tháng 1 năm 2007). “A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone”. Historical Biology. 19 (1): 23–33. doi:10.1080/08912960600845767. ISSN 0891-2963. S2CID 85819339.
- ^ Langer, Max C.; Ferigolo, Jorge (1 tháng 1 năm 2013). “The Late Triassic dinosauromorph Sacisaurus agudoensis (Caturrita Formation; Rio Grande do Sul, Brazil): anatomy and affinities”. Geological Society, London, Special Publications (bằng tiếng Anh). 379 (1): 353–392. Bibcode:2013GSLSP.379..353L. doi:10.1144/SP379.16. ISSN 0305-8719. S2CID 131414332.
- ^ Cabreira, S.F.; Kellner, A.W.A.; Dias-da-Silva, S.; da Silva, L.R.; Bronzati, M.; de Almeida Marsola, J.C.; Müller, R.T.; de Souza Bittencourt, J.; Batista, B.J.; Raugust, T.; Carrilho, R.; Brodt, A.; Langer, M.C. (2016). “A Unique Late Triassic Dinosauromorph Assemblage Reveals Dinosaur Ancestral Anatomy and Diet”. Current Biology. 26 (22): 3090–3095. doi:10.1016/j.cub.2016.09.040. PMID 27839975.
- ^ Müller, Rodrigo Temp; Garcia, Maurício Silva (26 tháng 8 năm 2020). “A paraphyletic 'Silesauridae' as an alternative hypothesis for the initial radiation of ornithischian dinosaurs”. Biology Letters. 16 (8): 20200417. doi:10.1098/rsbl.2020.0417. PMC 7480155. PMID 32842895.
- ^ “Từ điển Hán Nôm”. https://hvdic.thivien.net. Liên kết ngoài trong
|website=(trợ giúp) - ^ a b Owen 1842, p.103: "The combination of such characters … will, it is presumed, be deemed sufficient ground for establishing a distinct tribe or sub-order of Saurian Reptiles, for which I would propose the name of Dinosauria*. (*Gr. δεινός, fearfully great; σαύρος, a lizard. …)
- ^ “Dinosauria”. Merriam-Webster Dictionary. Truy cập ngày 10 tháng 11 năm 2019.
- ^ Crane, George R. (biên tập). “Greek Dictionary Headword Search Results”. Perseus 4.0. Medford và Somerville, MA: Tufts University. Truy cập ngày 13 tháng 10 năm 2019. Lemma for 'δεινός' from Henry George Liddell, Robert Scott, A Greek-English Lexicon (1940): 'fearful, terrible'.
- ^ Farlow & Brett-Surman 1997, tr. ix–xi, Preface, "Dinosaurs: The Terrestrial Superlative" by James O. Farlow and M.K. Brett-Surman.
- ^ a b Chamary, JV (ngày 30 tháng 9 năm 2014). “Dinosaurs, Pterosaurs And Other Saurs – Big Differences”. Forbes.com. Jersey City, NJ: Forbes Media, LLC. ISSN 0015-6914. Lưu trữ bản gốc ngày 10 tháng 11 năm 2014. Truy cập ngày 2 tháng 10 năm 2018.
- ^ a b c d Benton, Michael J. (2004). “Origin and relationships of Dinosauria”. Trong Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (biên tập). The Dinosauria (ấn bản thứ 2). Berkeley: University of California Press. tr. 7–19. ISBN 0-520-24209-2.Quản lý CS1: nhiều tên: danh sách biên tập viên (liên kết)
- ^ Olshevsky 2000
- ^ Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênLetal05 - ^ “Using the tree for classification”. Understanding Evolution. Berkeley: Đại học California. Lưu trữ bản gốc ngày 31 tháng 8 năm 2019. Truy cập ngày 14 tháng 10 năm 2019.
- ^ a b Weishampel, Dodson & Osmólska 2004, tr. 210–231, chpt. 11: "Basal Avialae" by Kevin Padian.
- ^ Wade, Nicholas (22 tháng 3 năm 2017). “Shaking Up the Dinosaur Family Tree”. The New York Times. New York. ISSN 0362-4331. Lưu trữ bản gốc ngày 7 tháng 4 năm 2018. Truy cập ngày 30 tháng 10 năm 2019. "A version of this article appears in print on March 28, 2017, on Page D6 of the New York edition with the headline: Shaking Up the Dinosaur Family Tree."
- ^ Baron, Matthew G.; Norman, David B.; Barrett, Paul M. (2017). “A new hypothesis of dinosaur relationships and early dinosaur evolution”. Nature. London: Nature Research. 543 (7646): 501–506. Bibcode:2017Natur.543..501B. doi:10.1038/nature21700. ISSN 0028-0836. PMID 28332513. S2CID 205254710. "This file contains Supplementary Text and Data, Supplementary Tables 1-3 and additional references.": Supplementary Information[liên kết hỏng]
- ^ Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênDFG97 - ^ Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênDL90 - ^ Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênMM97 - ^ Tennant, Jonathan P.; Chiarenza, Alfio Alessandro; Baron, Matthew (19 tháng 2 năm 2018). “How has our knowledge of dinosaur diversity through geologic time changed through research history?”. PeerJ. 6: e4417. doi:10.7717/peerj.4417. PMC 5822849. PMID 29479504.
- ^ Starrfelt, Jostein; Liow, Lee Hsiang (2016). “How many dinosaur species were there? Fossil bias and true richness estimated using a Poisson sampling model”. Philosophical Transactions of the Royal Society B: Biological Sciences. 371 (1691): 20150219. doi:10.1098/rstb.2015.0219. PMC 4810813. PMID 26977060.
- ^ Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênWang&Dodson - ^ Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênrussell1995 - ^ Starrfelt, Jostein; Liow, Lee Hsiang (2016). “How many dinosaur species were there? Fossil bias and true richness estimated using a Poisson sampling model”. Philosophical Transactions of the Royal Society B. London: Royal Society. 371 (1691): 20150219. doi:10.1098/rstb.2015.0219. ISSN 0962-8436. PMC 4810813. PMID 26977060.
- ^ Black, Riley (23 tháng 3 năm 2016). “Most Dinosaur Species Are Still Undiscovered”. National Geographic News. Lưu trữ bản gốc ngày 6 tháng 3 năm 2021. Truy cập ngày 6 tháng 6 năm 2021.
- ^ Gill, F.; Donsker, D.; Rasmussen, P. (2021). “Welcome”. IOC World Bird List 11.1.
- ^ a b MacLeod, Norman; Rawson, Peter F.; Forey, Peter L.; và đồng nghiệp (1997). “The Cretaceous–Tertiary biotic transition”. Journal of the Geological Society. London: Hội Địa chất Luân Đôn. 154 (2): 265–292. Bibcode:1997JGSoc.154..265M. doi:10.1144/gsjgs.154.2.0265. ISSN 0016-7649. S2CID 129654916.
- ^ a b Amiot, R.; Buffetaut, E.; Lecuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, M.A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; Zhou, Z. (2010). “Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods”. Geology. 38 (2): 139–142. Bibcode:2010Geo....38..139A. doi:10.1130/G30402.1.
- ^ a b c Brusatte, Stephen L. (2012). Dinosaur Paleobiology (ấn bản thứ 1). New York: Wiley, J. tr. 9–20, 21. ISBN 978-0-470-65658-7.
- ^ Nesbitt, Sterling J. (ngày 29 tháng 4 năm 2011). “The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades” (PDF). Bulletin of the American Museum of Natural History. New York: Bảo tàng Lịch sử Tự nhiên Hoa Kỳ. 2011 (352): 1–292. doi:10.1206/352.1. hdl:2246/6112. ISSN 0003-0090. Lưu trữ bản gốc ngày 29 tháng 2 năm 2016. Truy cập ngày 16 tháng 10 năm 2019.
- ^ Paul 2000, tr. 140–168, chpt. 3: "Classification and Evolution of the Dinosaur Groups" by Thomas R. Holtz Jr.
- ^ Smith, Dave. “Dinosauria Morphology”. UC Berkeley. Truy cập ngày 21 tháng 1 năm 2013.
- ^ Langer, Max C.; Abdala, Fernando; Richter, Martha; Benton, Michael J. (ngày 15 tháng 10 năm 1999). “Un dinosaure sauropodomorphe dans le Trias supérieur (Carnien) du Sud du Brésil” [A sauropodomorph dinosaur from the Upper Triassic (Carman) of southern Brazil]. Comptes Rendus de l'Académie des Sciences, Série IIA. Amsterdam: Elsevier on behalf of the French Academy of Sciences. 329 (7): 511–517. Bibcode:1999CRASE.329..511L. doi:10.1016/S1251-8050(00)80025-7. ISSN 1251-8050.
- ^ Nesbitt, Sterling J.; Irmis, Randall B.; Parker, William G. (2007). “A critical re-evaluation of the Late Triassic dinosaur taxa of North America”. Journal of Systematic Palaeontology. Milton Park, Oxfordshire: Taylor & Francis on behalf of the Natural History Museum, London. 5 (2): 209–243. doi:10.1017/S1477201907002040. ISSN 1477-2019.
- ^ This was recognized not later than 1909: Celeskey, Matt (2005). “Dr. W. J. Holland and the Sprawling Sauropods”. The Hairy Museum of Natural History. Bản gốc lưu trữ ngày 12 tháng 6 năm 2011. Truy cập ngày 18 tháng 10 năm 2019.
- Holland, William J. (tháng 5 năm 1910). “A Review of Some Recent Criticisms of the Restorations of Sauropod Dinosaurs Existing in the Museums of the United States, with Special Reference to that of Diplodocus Carnegiei in the Carnegie Museum”. The American Naturalist. American Society of Naturalists. 44 (521): 259–283. doi:10.1086/279138. ISSN 0003-0147. Truy cập ngày 18 tháng 10 năm 2019.
- The arguments and many of the images are also presented in Desmond 1975.
- ^ a b Benton 2005
- ^ Cowen 2005, tr. 151–175, chpt. 12: "Dinosaurs".
- ^ a b Kubo, Tai; Benton, Michael J. (tháng 11 năm 2007). “Evolution of hindlimb posture in archosaurs: limb stresses in extinct vertebrates”. Palaeontology. Hoboken, NJ: Wiley-Blackwell. 50 (6): 1519–1529. doi:10.1111/j.1475-4983.2007.00723.x. ISSN 0031-0239.
- ^ Kump LR, Pavlov A & Arthur MA (2005). “Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia”. Geology. 33 (5): 397–400. Bibcode:2005Geo....33..397K. doi:10.1130/G21295.1.
- ^ Tanner LH, Lucas SG & Chapman MG (2004). “Assessing the record and causes of Late Triassic extinctions” (PDF). Earth-Science Reviews. 65 (1–2): 103–139. Bibcode:2004ESRv...65..103T. doi:10.1016/S0012-8252(03)00082-5. Bản gốc (PDF) lưu trữ ngày 25 tháng 10 năm 2007. Truy cập ngày 22 tháng 10 năm 2007.
- ^ Sereno PC (1999). “The evolution of dinosaurs”. Science. 284 (5423): 2137–2147. doi:10.1126/science.284.5423.2137. PMID 10381873.
- ^ Sereno, P.C.; Forster, Catherine A.; Rogers, Raymond R.; Monetta, Alfredo M. (1993). “Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria”. Nature. 361 (6407): 64–66. Bibcode:1993Natur.361...64S. doi:10.1038/361064a0.
- ^ Nesbitt, S. J., Barrett, P. M., Werning, S., Sidor, C. A., and A. J. Charig. (2012). "The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania." Biology Letters.
- ^ a b c d Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênHCL04 - ^ a b c d e Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênFS04 - ^ Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênserenoetal07 - ^ Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênPSAS05 - ^ Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênAF04 - ^ Dyke & Kaiser 2011, chpt. 14: "Bird Evolution Across the K–Pg Boundary and the Basal Neornithine Diversification" by Bent E. K. Lindow. doi:10.1002/9781119990475.ch14
- ^ Cracraft, Joel (ngày 21 tháng 6 năm 1968). “A Review of the Bathornithidae (Aves, Gruiformes), with Remarks on the Relationships of the Suborder Cariamae” (PDF). American Museum Novitates. New York: American Museum of Natural History. 2326: 1–46. hdl:2246/2536. ISSN 0003-0082. Truy cập ngày 22 tháng 10 năm 2019.
- ^ Alvarenga, Herculano; Jones, Washington W.; Rinderknecht, Andrés (tháng 5 năm 2010). “The youngest record of phorusrhacid birds (Aves, Phorusrhacidae) from the late Pleistocene of Uruguay”. Neues Jahrbuch für Geologie und Paläontologie. Stuttgart: E. Schweizerbart. 256 (2): 229–234. doi:10.1127/0077-7749/2010/0052. ISSN 0077-7749. Truy cập ngày 22 tháng 10 năm 2019.
- ^ Mayr 2009
- ^ Paul 1988, tr. 248–250
- ^ Taylor, M.P.; Wedel, M.J. (2013). “Why sauropods had long necks; and why giraffes have short necks”. PeerJ. 1: e36. doi:10.7717/peerj.36. PMC 3628838. PMID 23638372.
- ^ Lỗi chú thích: Thẻ
<ref>sai; không có nội dung trong thẻ ref có tênNYT-20161208 - ^ Sander, P. Martin; Christian, Andreas; Clauss, Marcus; và đồng nghiệp (2011). “Biology of the sauropod dinosaurs: the evolution of gigantism”. Biological Reviews. 86 (1): 117–155. doi:10.1111/j.1469-185X.2010.00137.x. PMC 3045712. PMID 21251189.
- ^ a b c Carpenter, Kenneth (2006). “Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus”. Trong Foster, John R.; and Lucas, Spencer G. (biên tập). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque: New Mexico Museum of Natural History and Science. tr. 131–138.Quản lý CS1: nhiều tên: danh sách biên tập viên (liên kết)
- ^ Paul, Gregory S. (2010). Princeton Field Guide to Dinosaurs. Princeton University Press. ISBN 978-0-691-13720-9.
- ^ Colbert, Edwin Harris (1971). Men and dinosaurs: the search in field and laboratory. Harmondsworth [Eng.]: Penguin. ISBN 0-14-021288-4.
- ^ Lucas, H., Hecket, H. & S. (2004). "Reappraisal of Seismosaurus, a Late Jurassic Sauropod". Proceeding, Annual Meeting of the Society of Paleontology 36 (5): 422.
- ^ Lovelace, David M. (2007). “Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny”. Arquivos do Museu Nacional. 65 (4): 527–544.
- ^ dal Sasso C, Maganuco S, Buffetaut E, Mendez MA (2006). “New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities” (PDF). Journal of Vertebrate Paleontology. 25 (4): 888–896. doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2. Bản gốc (PDF) lưu trữ ngày 29 tháng 4 năm 2011. Truy cập ngày 5 tháng 5 năm 2011.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Therrien, F.; Henderson, D.M. (2007). “My theropod is bigger than yours... or not: estimating body size from skull length in theropods”. Journal of Vertebrate Paleontology. 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- ^ a b Zhang F, Zhou Z, Xu X, Wang X, Sullivan C (2008). “A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers”. Nature. 455 (7216): 1105–1108. Bibcode:2008Natur.455.1105Z. doi:10.1038/nature07447. PMID 18948955.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ a b Xu X, Zhao Q, Norell M, Sullivan C, Hone D, Erickson G, Wang XL, Han FL, Guo Y (2008). “A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin”. Chinese Science Bulletin. 54 (3): 430–435. doi:10.1007/s11434-009-0009-6.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Butler, R.J.; Zhao, Q. (2009). “The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus yansiensis from the Late Cretaceous of China”. Cretaceous Research. 30 (1): 63–77. doi:10.1016/j.cretres.2008.03.002.
- ^ doi:10.1098/rspb.2006.3532
Hoàn thành chú thích này - ^ Yans J, Dejax J, Pons D, Dupuis C & Taquet P (2005). “Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique)”. Comptes Rendus Palevol (bằng tiếng Pháp). 4 (1–2): 135–150. doi:10.1016/j.crpv.2004.12.003.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Day, J.J.; Upchurch, P; Norman, DB; Gale, AS; Powell, HP (2002). “Sauropod trackways, evolution, and behavior”. Science. 296 (5573): 1659. doi:10.1126/science.1070167. PMID 12040187.
- ^ Wright, Joanna L. (2005). “Steps in understanding sauropod biology”. Trong Curry Rogers, Kristina A.; and Wilson, Jeffrey A. (biên tập). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. tr. 252–284. ISBN 0-520-24623-3.Quản lý CS1: nhiều tên: danh sách biên tập viên (liên kết)
- ^ Varricchio, D.J.; Sereno, Paul C.; Xijin, Zhao; Lin, Tan; Wilson, Jeffery A.; Lyon, Gabrielle H. (2008). “Mud-trapped herd captures evidence of distinctive dinosaur sociality” (PDF). Acta Palaeontologica Polonica. 53 (4): 567–578. doi:10.4202/app.2008.0402. Truy cập ngày 6 tháng 5 năm 2011.
- ^ Lessem, Don; Glut, Donald F. (1993). “Allosaurus”. The Dinosaur Society's Dinosaur Encyclopedia. Random House. tr. 19–20. ISBN 0-679-41770-2.
- ^ Maxwell, W. D.; Ostrom, John (1995). “Taphonomy and paleobiological implications of Tenontosaurus–Deinonychus associations”. Journal of Vertebrate Paleontology. 15 (4): 707–712. doi:10.1080/02724634.1995.10011256.
- ^ “The Fighting Dinosaurs”. American Museum of Natural History. Bản gốc lưu trữ ngày 18 tháng 1 năm 2012. Truy cập ngày 5 tháng 12 năm 2007.
- ^ Carpenter, K. (1998). “Evidence of predatory behavior by theropod dinosaurs”. Gaia. 15: 135–144.[liên kết hỏng]
- ^ Rogers, Raymond R.; Krause, DW; Curry Rogers, K (2007). “Cannibalism in the Madagascan dinosaur Majungatholus atopus”. Nature. 422 (6931): 515–518. doi:10.1038/nature01532. PMID 12673249.
- ^ Schmitz, L.; Motani, R. (2011). “Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology”. Science. 332 (6030): 705–708. Bibcode:2011Sci...332..705S. doi:10.1126/science.1200043. PMID 21493820.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Varricchio DJ, Martin, AJ and Katsura, Y (2007). “First trace and body fossil evidence of a burrowing, denning dinosaur”. Proceedings of the Royal Society B. 274 (1616): 1361–1368. doi:10.1098/rspb.2006.0443. PMC 2176205. PMID 17374596.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Chiappe, L.M. and Witmer, L.M. (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. ISBN 0-520-20094-2
- ^ Chatterjee, S.; Templin, R. J. (2007). “Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui” (PDF). Proceedings of the National Academy of Sciences. 104 (5): 1576–1580. Bibcode:2007PNAS..104.1576C. doi:10.1073/pnas.0609975104. PMC 1780066. PMID 17242354. Bản gốc (PDF) lưu trữ ngày 13 tháng 10 năm 2008. Truy cập ngày 28 tháng 9 năm 2014.
- ^ Currie, Philip J and Kevin Padian biên tập (1997). Encyclopedia of Dinosaurs. Academic Press. tr. 206.
|title=trống hay bị thiếu (trợ giúp) - ^ Hansell M (2000). Bird Nests and Construction Behaviour. University of Cambridge Press ISBN 0-521-46038-7
- ^ Varricchio, David J.; Horner, John J.; Jackson, Frankie D. (2002). “Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus”. Journal of Vertebrate Paleontology. 22 (3): 564–576. doi:10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2.
- ^ Dingus, L. and Rowe, T. (1998). The Mistaken Extinction – Dinosaur Evolution and the Origin of Birds. New York: W. H. Freeman.
- ^ doi:10.1016/j.epsl.2008.01.015
Hoàn thành chú thích này - ^ Mullen, L. (2004). "Multiple Impacts". Astrobiology Magazine.
- ^ doi:10.1144/gsjgs.154.2.0265
Hoàn thành chú thích này - ^ Miller KG, Kominz MA, Browning JV, Wright JD, Mountain GS, Katz ME, Sugarman PJ, Cramer BS, Christie-Blick N, Pekar SF (2005). “The Phanerozoic record of global sea-level change”. Science. 310 (5752): 1293–8. Bibcode:2005Sci...310.1293M. doi:10.1126/science.1116412. PMID 16311326.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ McArthura JM, Janssenb NMM, Rebouletc S, Lengd MJ, Thirlwalle MF & van de Shootbruggef B (2007). “Palaeotemperatures, polar ice-volume, and isotope stratigraphy (Mg/Ca, δ18O, δ13C, 87Sr/86Sr): The Early Cretaceous (Berriasian, Valanginian, Hauterivian)”. Palaeogeography, Palaeoclimatology, Palaeoecology. 248 (3–4): 391–430. doi:10.1016/j.palaeo.2006.12.015.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Paul, Gregory S. (2002). Dinosaurs of the air: the evolution and loss of flight in dinosaurs and birds. Johns Hopkins University Press. tr. 397. ISBN 0-8018-6763-0.
- ^ doi:10.1029/97JE01743
Hoàn thành chú thích này - ^ Alvarez, LW, Alvarez, W, Asaro, F, and Michel, HV (1980). “Extraterrestrial cause for the Cretaceous–Tertiary extinction”. Science. 208 (4448): 1095–1108. Bibcode:1980Sci...208.1095A. doi:10.1126/science.208.4448.1095. PMID 17783054.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Hildebrand, Alan R.; Penfield, Glen T.; Kring, David A.; Pilkington, Mark; Zanoguera, Antonio Camargo; Jacobsen, Stein B.; Boynton, William V. (tháng 9 năm 1991). “Chicxulub Crater; a possible Cretaceous/Tertiary boundary impact crater on the Yucatan Peninsula, Mexico”. Geology. 19 (9): 867–871. Bibcode:1991Geo....19..867H. doi:10.1130/0091-7613(1991)019<0867:CCAPCT>2.3.CO;2.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Pope KO, Ocampo AC, Kinsland GL, Smith R (1996). “Surface expression of the Chicxulub crater”. Geology. 24 (6): 527–30. Bibcode:1996Geo....24..527P. doi:10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2. PMID 11539331.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Keller G, Adatte T, Gardin S, Bartolini A, Bajpai S (2008). “Main Deccan volcanism phase ends near the K–T boundary: Evidence from the Krishna-Godavari Basin, SE India”. Earth and Planetary Science Letters. 268 (3–4): 293–311. Bibcode:2008E&PSL.268..293K. doi:10.1016/j.epsl.2008.01.015.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Duncan RA, Pyle DG (1988). “Rapid eruption of the Deccan flood basalts at the Cretaceous/Tertiary boundary”. Nature. 333 (6176): 841–843. Bibcode:1988Natur.333..841D. doi:10.1038/333841a0.
- ^ Alvarez, W (1997). T. rex and the Crater of Doom. Princeton University Press. tr. 130–146. ISBN 978-0-691-01630-6.
- ^ Fassett, JE, Lucas, SG, Zielinski, RA, and Budahn, JR; Lucas; Zielinski; Budahn (2001). “Compelling new evidence for Paleocene dinosaurs in the Ojo Alamo Sandstone, San Juan Basin, New Mexico and Colorado, USA” (PDF). Catastrophic events and mass extinctions, Lunar and Planetary Contribution. 1053: 45–46. Bibcode:2001caev.conf.3139F. Truy cập ngày 18 tháng 5 năm 2007.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Sloan, R. E., Rigby, K,. Van Valen, L. M., Gabriel, Diane (1986). “Gradual dinosaur extinction and simultaneous ungulate radiation in the Hell Creek Formation”. Science. 232 (4750): 629–633. Bibcode:1986Sci...232..629S. doi:10.1126/science.232.4750.629. PMID 17781415.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
Thư mục
[sửa | sửa mã nguồn]- Alvarez, Walter (1997). T. rex and the Crater of Doom. Princeton, NJ: Princeton University Press. ISBN 978-0-691-01630-6. LCCN 96049208. OCLC 1007846558. Truy cập ngày 4 tháng 11 năm 2019.
- Bakker, Robert T. (1986). The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and Their Extinction. New York: William Morrow and Company. ISBN 978-0-688-04287-5. LCCN 86012643. OCLC 13699558. Truy cập ngày 6 tháng 11 năm 2019.
- Benton, Michael J. (2005). Vertebrate Palaeontology (ấn bản thứ 3). Malden, MA: Blackwell Publishing. ISBN 978-0-632-05637-8. LCCN 2003028152. OCLC 53970617. Truy cập ngày 30 tháng 10 năm 2019.
- Brusatte, Stephen L. (2012). Benton, Michael J. (biên tập). Dinosaur Paleobiology. Topics in Paleobiology. Foreword by Michael J. Benton. Hoboken, NJ: Wiley-Blackwell. doi:10.1002/9781118274071. ISBN 978-0-470-65658-7. LCCN 2011050466. OCLC 781864955.
- Chiappe, Luis M.; Witmer, Lawrence M. biên tập (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. ISBN 978-0-520-20094-4. LCCN 2001044600. OCLC 901747962.
- Colbert, Edwin H. (1971) [Originally published, New York: E. P. Dutton, 1968; London: Evans Brothers Ltd, 1969]. Men and Dinosaurs: The Search in Field and Laboratory. Harmondsworth: Penguin. ISBN 978-0-14-021288-4. OCLC 16208760. Truy cập ngày 31 tháng 10 năm 2019.
- Cowen, Richard (2005). History of Life (ấn bản thứ 4). Malden, MA: Blackwell Publishing. ISBN 978-1-4051-1756-2. LCCN 2003027993. OCLC 53970577. The 5th edition of the book is available from the Internet Archive. Retrieved 2019-10-19.
- Currie, Philip J.; Padian, Kevin biên tập (1997). Encyclopedia of Dinosaurs. San Diego, CA: Academic Press. ISBN 978-0-12-226810-6. LCCN 97023430. OCLC 436848919. Truy cập ngày 30 tháng 10 năm 2019.
- Currie, Philip J.; Koppelhus, Eva B.; Shugar, Martin A.; Wright, Joanna L. biên tập (2004). Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Life of the Past. Bloomington, IN: Indiana University Press. ISBN 978-0-253-34373-4. LCCN 2003019035. OCLC 52942941.
- Curry Rogers, Kristina A.; Wilson, Jeffrey A. biên tập (2005). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. ISBN 978-0-520-24623-2. LCCN 2005010624. OCLC 879179542.
- Desmond, Adrian J. (1975). The Hot-Blooded Dinosaurs: A Revolution in Palaeontology. London: Blond & Briggs. ISBN 978-0-8037-3755-6. LCCN 76359907. OL 4933052M. Truy cập ngày 30 tháng 10 năm 2019.
- Dickens, Charles (1853). Bleak House. London: Bradbury and Evans. Truy cập ngày 7 tháng 11 năm 2019.
- Dodson, Peter; Gingerich, Philip D. biên tập (1993). “Functional Morphology and Evolution”. The American Journal of Science and Arts. A special volume of the American Journal of Science. New Haven, CT: Kline Geology Laboratory, Yale University. 293-A. ISSN 0002-9599. OCLC 27781160.
- Dong, Zhiming (1992). Dinosaurian Faunas of China . Beijing; Berlin; New York: China Ocean Press; Springer-Verlag. ISBN 978-3-540-52084-9. LCCN 92207835. OCLC 26522845.
- Dyke, Gareth; Kaiser, Gary biên tập (2011). Living Dinosaurs: The Evolutionary History of Modern Birds. Chichester; Hoboken, NJ: Wiley-Blackwell. ISBN 978-0-470-65666-2. LCCN 2010043277. OCLC 729724640.
- Farlow, James O.; Brett-Surman, M.K. biên tập (1997). The Complete Dinosaur. Bloomington, IN: Indiana University Press. ISBN 978-0-253-33349-0. LCCN 97-23698. OCLC 924985811. Truy cập ngày 14 tháng 10 năm 2019.
- Foster, John R.; Lucas, Spencer G. biên tập (2006). “Paleontology and Geology of the Upper Jurassic Morrison Formation”. Bulletin of the New Mexico Museum of Natural History and Science. New Mexico Museum of Natural History and Science Bulletin. Albuquerque, NM: New Mexico Museum of Natural History and Science. 36. ISSN 1524-4156. OCLC 77520577. Truy cập ngày 21 tháng 10 năm 2019.
- Glut, Donald F. (1997). Dinosaurs: The Encyclopedia. Foreword by Michael K. Brett-Surman. Jefferson, NC: McFarland & Company. ISBN 978-0-89950-917-4. LCCN 95047668. OCLC 33665881.
- Gunther, Robert Theodore biên tập (1968) [First printed in Oxford 1945]. Life and Letters of Edward Lhwyd. Early Science in Oxford. XIV. Preface by Albert Everard Gunther . London: Dawsons of Pall Mall. ISBN 978-0-7129-0292-2. LCCN 22005926. OCLC 43529321. Truy cập ngày 4 tháng 11 năm 2019.
- Hansell, Mike (2000). Bird Nests and Construction Behaviour. Pen and ink illustration by Raith Overhill. Cambridge: University of Cambridge Press. ISBN 978-0-521-46038-5. LCCN 99087681. OCLC 876286627. Truy cập ngày 30 tháng 10 năm 2019.
- Heilmann, Gerhard (1926). The Origin of Birds. London; New York: H. F. & G. Witherby; D. Appleton & Company. LCCN 27001127. OCLC 606021642.
- Holmes, Thom (1998). Fossil Feud: The Rivalry of the First American Dinosaur Hunters. Parsippany, NJ: Julian Messner. ISBN 978-0-382-39149-1. LCCN 96013610. OCLC 34472600.
- Holtz, Thomas R. Jr. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Illustrated by Luis V. Rey. New York: Random House. ISBN 978-0-375-82419-7. LCCN 2006102491. OCLC 77486015. Truy cập ngày 22 tháng 10 năm 2019.
- Lambert, David; The Diagram Group (1990). The Dinosaur Data Book: The Definitive, Fully Illustrated Encyclopedia of Dinosaurs. New York: Avon Books. ISBN 978-0-380-75896-8. LCCN 89092487. OCLC 21833417. Truy cập ngày 14 tháng 10 năm 2019.
- Lessem, Don; Glut, Donald F. (1993). The Dinosaur Society's Dinosaur Encyclopedia. Illustrations by Tracy Lee Ford; scientific advisors, Peter Dodson, et al. New York: Random House. ISBN 978-0-679-41770-5. LCCN 94117716. OCLC 30361459. Truy cập ngày 30 tháng 10 năm 2019.
- Lhuyd, Edward (1699). Lithophylacii Britannici ichnographia [British figured stones]. London: Ex Officina M.C. Bản gốc lưu trữ ngày 14 tháng 8 năm 2020. Truy cập ngày 4 tháng 11 năm 2019.
- Mayr, Gerald (2009). Paleogene Fossil Birds. Berlin: Springer-Verlag. doi:10.1007/978-3-540-89628-9. ISBN 978-3-540-89627-2. LCCN 2008940962. OCLC 916182693. S2CID 88941254. Truy cập ngày 30 tháng 10 năm 2019.
- Norell, Mark; Gaffney, Eugene S.; Dingus, Lowell (2000) [Originally published as Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf, 1995]. Discovering Dinosaurs: Evolution, Extinction, and the Lessons of Prehistory . Berkeley: University of California Press. ISBN 978-0-520-22501-5. LCCN 99053335. OCLC 977125867. Truy cập ngày 30 tháng 10 năm 2019.
- Olshevsky, George (2000). An Annotated Checklist of Dinosaur Species by Continent. Mesozoic Meanderings. 3. Illustrated by Tracy Lee Ford. San Diego, CA: Publications Requiring Research. ISSN 0271-9428. LCCN 00708700. OCLC 44433611.
- Owen, Richard (1842). “Report on British Fossil Reptiles. Part II”. Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841. London: John Murray. tr. 60–204. ISBN 978-0-8201-1526-9. LCCN 99030427. OCLC 1015526268. Truy cập ngày 13 tháng 10 năm 2019.
- Padian, Kevin biên tập (1986). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences. 8. San Francisco, CA: California Academy of Sciences. ISBN 978-0-940228-14-6. OCLC 946083441. OL 9826926M.
- Parsons, Keith M. (2001). Drawing out Leviathan: Dinosaurs and the Science Wars. Life in the Past. Bloomington, IN: Indiana University Press. ISBN 978-0-253-33937-9. LCCN 2001016803. OCLC 50174737. Truy cập ngày 30 tháng 10 năm 2019.
- Paul, Gregory S. (1988). Predatory Dinosaurs of the World: A Complete Illustrated Guide. New York: Simon & Schuster. ISBN 978-0-671-61946-6. LCCN 88023052. OCLC 859819093. Truy cập ngày 30 tháng 10 năm 2019.
- Paul, Gregory S. biên tập (2000). The Scientific American Book of Dinosaurs (ấn bản thứ 1). New York: St. Martin's Press. ISBN 978-0-312-26226-6. LCCN 2001269051. OCLC 45256074.
- Paul, Gregory S. (2010). The Princeton Field Guide to Dinosaurs. Princeton Field Guides. Princeton, NJ: Princeton University Press. ISBN 978-0-691-13720-9. LCCN 2010014916. OCLC 907619291.
- Plot, Robert (1677). The Natural History of Oxford-shire: Being an Essay toward the Natural History of England. Printed at the Theater in OXFORD, and are to be had there: And in London at Mr. S. Millers, at the Star near the West-end of St. Pauls Church-yard. Oxford; London. LCCN 11004267. OCLC 933062622. Truy cập ngày 13 tháng 11 năm 2019.
- Randall, Lisa (2015). Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the Universe. New York: HarperCollins: Ecco. ISBN 978-0-06-232847-2. LCCN 2016427646. OCLC 962371431.
- Rupke, Nicolaas A. (1994). Richard Owen: Victorian Naturalist. New Haven: Yale University Press. ISBN 978-0-300-05820-8. LCCN 93005739. OCLC 844183804. Truy cập ngày 5 tháng 11 năm 2019.
- Sarjeant, William A.S. biên tập (1995). Vertebrate Fossils and the Evolution of Scientific Concepts: Writings in Tribute to Beverly Halstead, by Some of His Many Friends. Modern Geology. Amsterdam: Gordon and Breach Publishers. ISBN 978-2-88124-996-9. ISSN 0026-7775. LCCN 00500382. OCLC 34672546. "Reprint of papers published in a special volume of Modern geology [v. 18 (Halstead memorial volume), 1993], with five additional contributions.--Pref."
- Tanner, Lawrence H.; Spielmann, Justin A.; Lucas, Spencer G. biên tập (2013). “The Triassic System: New Developments in Stratigraphy and Paleontology”. Bulletin of the New Mexico Museum of Natural History and Science. New Mexico Museum of Natural History and Science Bulletin. Albuquerque, NM: New Mexico Museum of Natural History and Science. 61. ISSN 1524-4156. OCLC 852432407. Truy cập ngày 21 tháng 10 năm 2019.
- Weishampel, David B.; Dodson, Peter; Osmólska, Halszka biên tập (2004). The Dinosauria (ấn bản thứ 2). Berkeley: University of California Press. ISBN 978-0-520-25408-4. LCCN 2004049804. OCLC 154697781.
Đọc thêm
[sửa | sửa mã nguồn]- University of Southampton (29 tháng 9 năm 2021). “Two New Species of Large Predatory Dinosaur With Crocodile-Like Skulls Discovered on Isle of Wight”. SciTechDaily.
- Zhou, Zhonghe (tháng 10 năm 2004). “The origin and early evolution of birds: discoveries, disputes, and perspectives from fossil evidence” (PDF). Naturwissenschaften. Berlin: Springer Science+Business Media. 91 (10): 455–471. Bibcode:2004NW.....91..455Z. doi:10.1007/s00114-004-0570-4. ISSN 0028-1042. PMID 15365634. S2CID 3329625. Bản gốc (PDF) lưu trữ ngày 21 tháng 7 năm 2011. Truy cập ngày 6 tháng 11 năm 2019.
- Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore; London: Johns Hopkins University Press. ISBN 978-0-8018-6763-7. LCCN 2001000242. OCLC 1088130487..
- Stewart, Tabori & Chang (1997). The Humongous Book of Dinosaurs. New York: Stewart, Tabori & Chang. ISBN 978-1-55670-596-0. LCCN 97000398. OCLC 1037269801.
- Sternberg, Charles Mortram (1966) [Original edition published by E. Cloutier, printer to the King, 1946]. Canadian Dinosaurs. Geological Series. 54 (ấn bản thứ 2). Ottawa: National Museum of Canada. LCCN gs46000214. OCLC 1032865683.
Liên kết ngoài
[sửa | sửa mã nguồn]| Thư viện tài nguyên ngoại văn về Dinosaurs |
| Từ điển từ Wiktionary | |
| Tập tin phương tiện từ Commons | |
| Danh ngôn từ Wikiquote | |
| Văn kiện từ Wikisource | |
| Tài nguyên học tập từ Wikiversity | |
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||









| |||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||


| Giải phẫu học | |||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hành vi | |||||||||||||||||||||||||||||||||||||||||||||||
| Tiến hóa | |||||||||||||||||||||||||||||||||||||||||||||||
| Hóa thạch | |||||||||||||||||||||||||||||||||||||||||||||||
| Tương tác với người | |||||||||||||||||||||||||||||||||||||||||||||||
| Danh sách | |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| Tiến hóa |
|
|---|---|
| Di truyền học quần thể | |
| Phát triển | |
| Của việc phân loại | |
| Của các cơ quan | |
| Của các quá trình | |
| Tempo và mode | |
| Sự hình thành loài | |
| Lịch sử | |
| Triết học | |
| Liên quan | |

.png){kind=link}
![[Tập hiểu sâu]: Vì sao Bạn luôn KHÔNG ỔN?](https://images.spiderum.com/sp-images/f5ab2630336211eea9e7ef608f381f46.png) GIẢM
12%
GIẢM
12%
 GIẢM
30%
GIẢM
30%
 GIẢM
50%
GIẢM
50%
 GIẢM
18%
GIẢM
18%
 GIẢM
50%
GIẢM
50%



